







130 milionów lat przed dniem dzisiejszym
![]()
Epoka: mezozoik / kreda
 Rozwój roślin kwitnących rozpoczął się prawdopodobnie około 200 milionów lat temu, kiedy niektóre rośliny zwielokrotniły cały swój materiał genetyczny. Dalsze zmiany w namnożonych chromosomach doprowadziły do powstania kwiatów. Rozwój ten musiał mieć miejsce kilka razy, o czym świadczy ilość podstawowych schematów budowy kwiatów.
Rozwój roślin kwitnących rozpoczął się prawdopodobnie około 200 milionów lat temu, kiedy niektóre rośliny zwielokrotniły cały swój materiał genetyczny. Dalsze zmiany w namnożonych chromosomach doprowadziły do powstania kwiatów. Rozwój ten musiał mieć miejsce kilka razy, o czym świadczy ilość podstawowych schematów budowy kwiatów.
Najstarsze skamieniałe rośliny kwitnące mają zaledwie około 130 milionów lat i występują bardzo rzadko. Stąd ich rozwój jest słabo zbadany. Być może rozwinęły się z paproci, które były identyfikowalne od 400 milionów lat. Rośliny kwitnące nadal mają wpływ na rozwój owadów i odwrotnie.
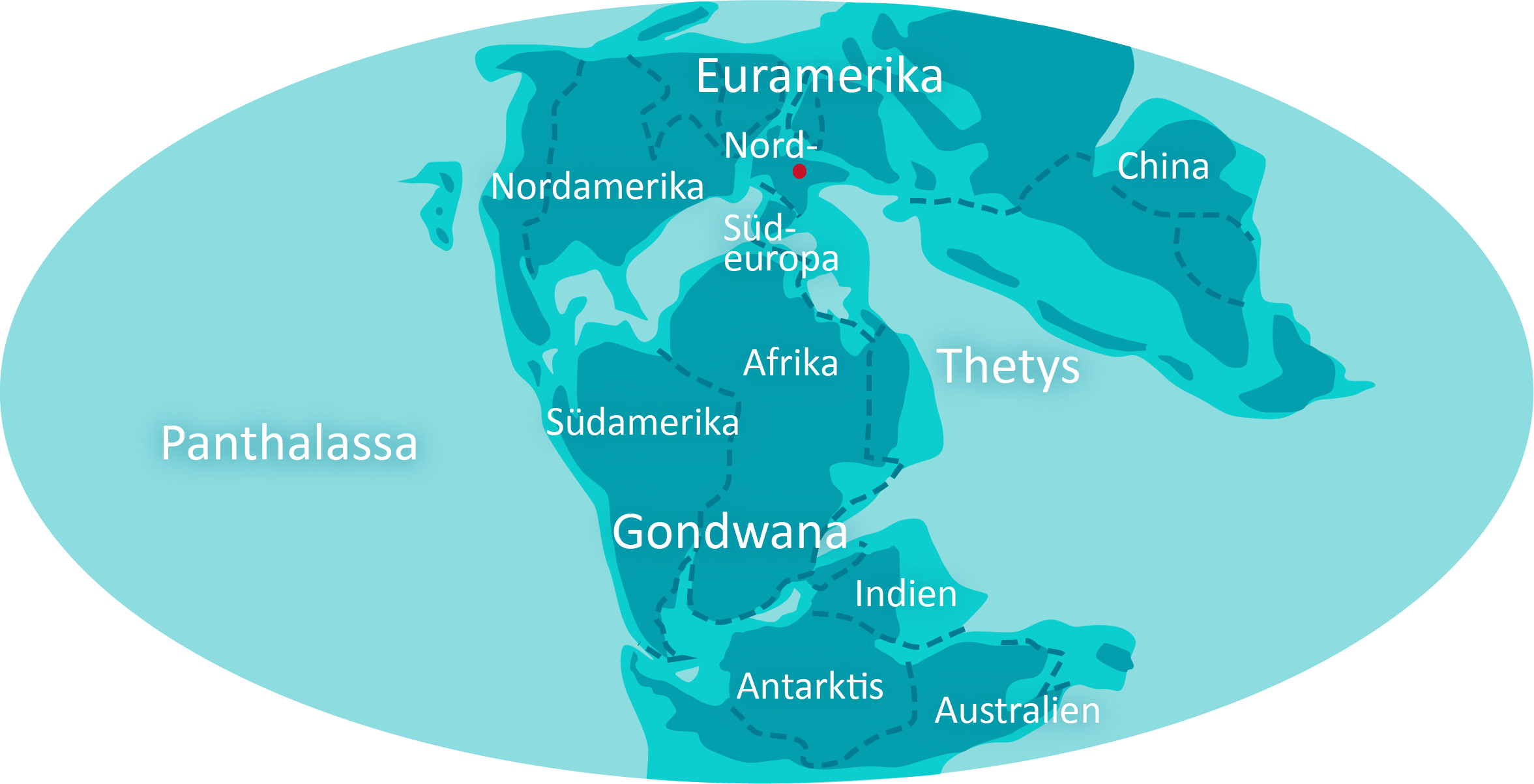 Klimat jest ciepły przez cały czas. Ląd jest wolny od lodu, więc poziom morza jest wysoki. Atmosfera zawiera 1,5 razy więcej tlenu i 4 razy więcej CO2 niż obecnie. Rozpad Gondwany trwa. Australia oddziela się od Antarktydy a Afryka od Ameryki Południowej, oddzielają się także Indie.
Klimat jest ciepły przez cały czas. Ląd jest wolny od lodu, więc poziom morza jest wysoki. Atmosfera zawiera 1,5 razy więcej tlenu i 4 razy więcej CO2 niż obecnie. Rozpad Gondwany trwa. Australia oddziela się od Antarktydy a Afryka od Ameryki Południowej, oddzielają się także Indie.
Tę informację w prostym języku znajdziesz na stronie evokids.de.
Grupa roślin nasiennych obejmuje praktycznie wszystkie żyjące współcześnie rośliny lądowe z wyjątkiem paproci i mchów. Pierwsze rośliny nasienne pojawiły się w dewonie. W karbonie rozwinęły się szkielety wspierające i umożlowiły rozwój większych form wzrostu.
Z reguły grupa okrytozalążkowych (Angiospermen) jest nazywana „roślinami kwiatowymi”. Ich dokładne pochodzenie jest nadal niejasne. Przypuszcza się, że podobnie jak grupa nagonasiennych (Gymnospermen), rozwinęły się one z grupy rdzennej paproci nasiennej. Jak sama nazwa wskazuje, łączy je wszystkie zdolność do produkcji nasion.
Najstarsze skamieniałości (pozostałości pyłku i liści) wskazują na pojawienie się pierwszych okrytozalążkowych już w jurze. W okresie kredy nastąpiła wręcz wybuchowa ewolucja roślin kwiatowych o wysokim wskaźniku specjacji.
Dominacja nagonasiennych
Do końca jury oprócz mchów i paproci oraz różnych pierwotnych form roślinnych, w tym żyjących do dziś sagowcow, w krajobrazie dominowały wspomniane już nagonasienne (Gymnospermen). Oprócz miłorzębu (ginko) jako „żywej skamieliny” obejmują one na przykład wszystkie żyjące do dziś drzewa iglaste, rodzinę cyprysów i gatunki jałowców. Ich zalążki leżą otwarcie na liściach zarodnionośnych (łuskach nasiennych).
Te z kolei są często połączone, tworząc okrągłe, ściśle ułożone kwiatostany na krótkich pędach. Nawiasem mówiąc, krótkie pędy i łuski nasienne kwiatów żeńskich są często zdrewniałe, co widać wyraźnie na przykładzie szyszek sosny.
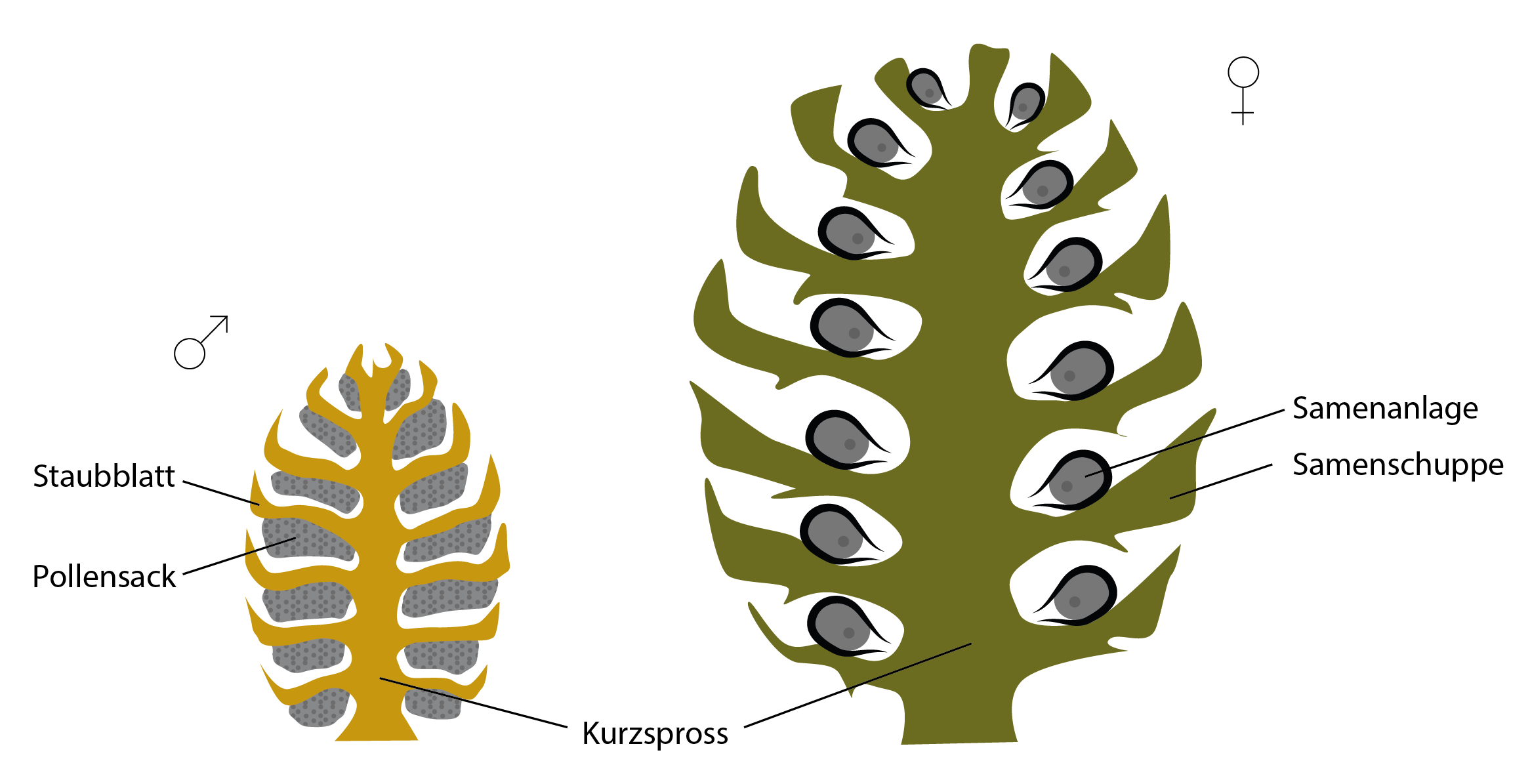
Kwiatostan nagonasiennych (po lewej męski, po prawej żeński). Zalążki kwiatostanu żeńskiego leżą odkryte na łusce nasiennej.

Kwiaty nagonasiennych. Po lewej: kwitnąca szyszka modrzewia; po prawej: szyszka świerkowa. (Źródła: https://commons.wikimedia.org/wiki/File:Kywiat.jpg, http://commons.wikimedia.org/wiki/File:Picea_Abies_female_cone_cropped.jpg)
{kind=link}
{kind=link}
Żeńskie kwiaty nagonasiennych są zwykle zapylane przez wiatr, czasami również z udziałem owadów (głównie chrząszczy).
Zwielokrotnione chromosomy — możliwości rozwoju
Już około 320 milionów lat temu doszło do zwielokrotnienia całego materiału genetycznego w prekursorach roślin nasiennych. Około 200 milionów lat temu takie zwielokrotnienie wystąpiło również u przodków dzisiejszych roślin kwitnących. Przez obszerne porównania grup genów dzisiejszych roślin kwitnących, mchów i widliczkowców, które wyłoniły się z pomnożonych genów, zawężono okresy tych zwielokrotnień.
Podczas gdy organizmy zwierzęce z wielokrotnie występującymi chromosomami (z kilkoma wyjątkami kilku pojedyńczych trisomii) na ogół nie są zdolne do życia, komórkom roślinnym nie wydają się sprawiać żadnych problemów. One nawet ciągną korzyści ze zwielokrotnionych chromosomów. Chociaż dużo z wielokrotnie występujących genów zniknęło z czasem, inne uległy przemianie się i przejęły nowe funkcje. Między innymi kontrolowały nowo powstałe procesy związane z kwitnieniem.
Pojawienie się okrytonasiennych
W okresie kredy rozwinęły się rośliny, które nie mają już otwarcie leżących zalążków, ale których owocnik jest zwinięty. Zaletą tego rozwiązania było to, że zalążki znajdujące się wewnątrz są chronione. Rośliny, u których ma to miejsce, określa się jako rośliny okrytozalążkowe (Angiospermen) lub po prostu „rośliny kwitnące”. W roślinach, które mają kilka owocolistków na kwiat, są one razem zrośnięte na różne sposoby, zawsze z tym skutkiem, że zalążki znajdują się chronione we wnętrzu. W ten sposób zwierzęta mogą służyć jako zapylacze bez wielkiego ryzyka zjedzenia zalążków.
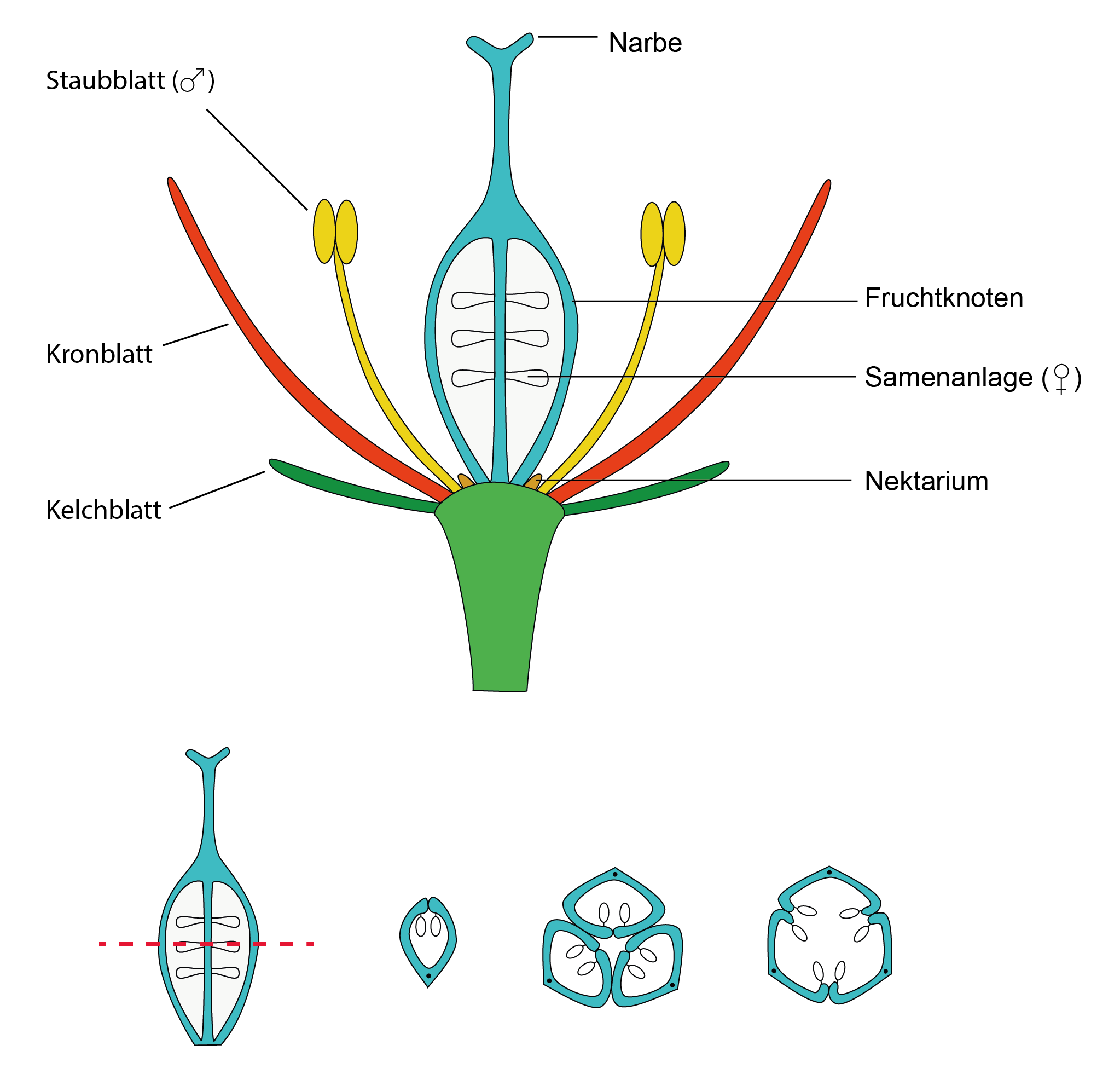
Budowa kwiatu rośliny okrytozalążkowej. Kwiat składa się ze wyspecjalizowanych liści. Płatki korony są często bardzo różnorodne. Pręciki wytwarzają (męski) pyłek, owocolistki niosą (żeńskie) zalążki i są zrośnięte, tworząc zalążnię. W dolnej części rysunku pokazano trzy z wielu możliwości budowy zalążni.
Pyłek nie trafia już bezpośrednio w zalążek, lecz w górną część zrośniętych owocolistków zwaną znamieniem. Stamtąd przez łagiewkę pyłkową dwa męskie plemniki pyłku docierają do żeńskiego zalążka i jego towarzyszy wewnątrz zalążni.
Konstrukcja ta oznacza, że tylko ten pyłek spośród wielu, które wylądują na znamieniu, może przeprowadzić zapłodnienie, który faktycznie jest w stanie jak najszybciej utworzyć stabilną łagiewkę pyłkową.
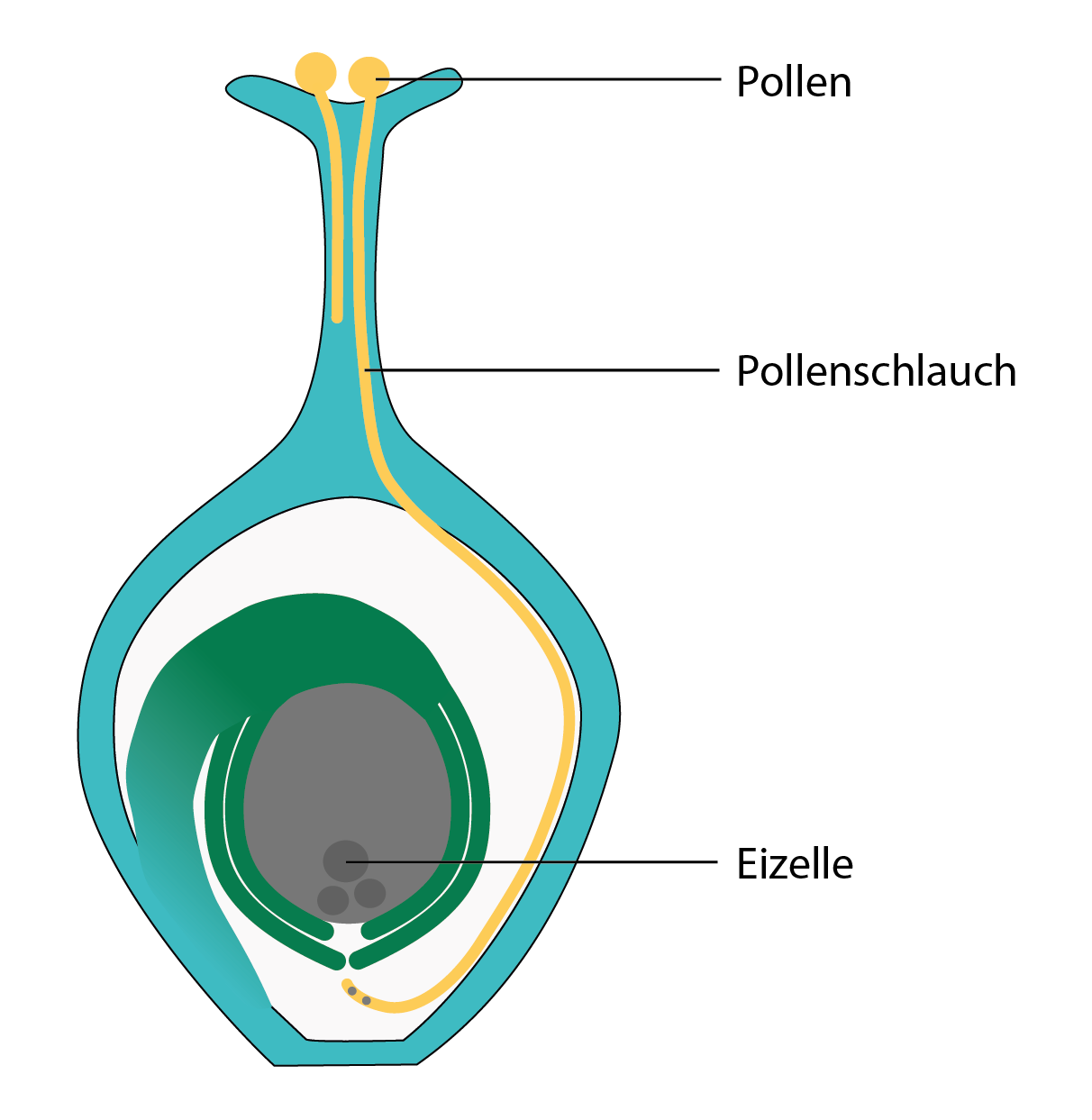
Łagiewka pyłkowa. Pyłek tworzy łagiewkę, przez którą jego jądra transportowane są do komórki jajowej. Żeński zalążek znajduje się wewnątrz zalężni i jest chroniony przez otaczające struktury.
Nie tylko obecność organów kwiatowych i ich budowa różni rośliny okrytozalążkowe od innych grup roślin. Do zróżnicowania przyczynia się również wiele innych cech, takich jak morfologia pyłku, charakterystyczne zróżnicowanie tkanki przewodzącej oraz różnorodność wtórnych substancji roślinnych, np. substancje zapachowe czy też służące ochronie przed roślinożercami (lektyny).
Tkanka odżywcza dla zarodka: ekonomia podwójnego zapłodnienia
Charakterystyczne jest również to, że w trakcie zapłodnienia u okrytozalążkowych oba jądra plemników przedostają się z łagiewki pyłkowej do zalążka podczas zapłodnienia (tzw. zapłodnienie podwójne). Jedno z nich zapładnia jajo żeńskie, a drugie, do zalężni wyposażonej już w dwa jądra, dodaje trzecie jądro. Ten triploidalny worek zarodkowy dzieli się i przekształca w tkankę odżywczą dla zarodka (tak zwane bielmo wtórne).
Z drugiej strony w przypadku nagonasiennych substancje magazynujące są często przechowywane w zalążku przed zapłodnieniem (np. orzeszki piniowe, orzeszki cedrowe). Ta haploidalna tkanka odżywcza nazywana jest bielmem pierwotnym.
Zaleta inwestowania energii w budowanie tkanki odżywczej po zapłodnieniu jest oczywista. Oprócz tego, że tkanka odżywcza nagozalążkowych jest zagrożona pożarciem, jest ona marnowana, jeśli nie nastąpi zapłodnienie. Z drugiej strony, jeśli jej powstawanie jest inicjowane tylko razem z zapłodnieniem, dzieje się to dokładnie wtedy, gdy jest to rzeczywiście potrzebne.
Koewolucja kwiatów, zapylaczy i dystrybutorów nasion
Oprócz rozwoju ochronnego woreczka zarodkowego, kwiaty okrytozalążkowców rozwinęły różne specjalizacje płatków, które przyciągają owady zapylające. Dalsze osiągnięcia, takie jak nektar i gruczoły zapachowe, są dalszymi zaletami.
W trakcie tego procesu rozwinęły się bardzo szczególne współzależności między owadami i roślinami. Na przykład niektóre owady koncentrują się na bardzo specyficznych gatunkach roślin, przez które są przyciągane, i z kolei rozwinęły odpowiadające im narządy (np. długa trąbka u motyli lub worki pyłkowe u pszczół), aby dostać się do substancji wabiących (atraktantów) roślin i dalej przy tej okazji, z korzyścią dla rośliny, zapewnić zapłodnienie.
Więcej informacji, a przede wszystkim ładne zdjęcia można znaleźć np. tutaj: http://deinnachbarlohbach.blogspot.com/2015/04/blutenbestauber.html
W toku ewolucji rozwinęła się również fascynująca współpraca między roślinami i zwierzętami w zakresie rozprzestrzeniania zapłodnionych nasion. Aby osiągnąć możliwie najszersze rozpowszechnienie, wiele roślin tworzy mięsiste owoce z części kwiatów wokół nasion. Są one zjadane przez zwierzęta, a nasiona są wydalane w odległym miejscu. W przypadku niektórych nasion przejście przez przewód pokarmowy rozprzestrzeniającego zwierzęcia zwiększa się prawdopodobieństwo, że w ogóle wykiełkują one pomyślnie.
Dwie duże grupy roślin kwiatowych
Rośliny okrytonasienne są tradycyjnie dzielone na dwie duże grupy na podstawie liczby ich liścieni:
- Tak zwane rośliny jednoliścienne po wykiełkowaniu tworzą pojedynczy liścień. Ich liście są zwykle wydłużone, żyłki liści są ułożone równolegle, a kwiaty mają potrójną symetrię (po 3 działki kielicha, płatki korony, pręciki i owocolistki lub ich wielokrotności). Przykładem są wszystkie trawy, ale także lilie i tulipany.
- Tak zwane dwuliścienne tworzą, jak sama nazwa wskazuje, dwa liścienie. Ich nerwy liściowe są rozgałęzione, a kwiaty często mają symetrię poczwórną (np. rzepak) lub popiątną (np. wszystkie drzewa owocowe).
Obecnie na całym świecie istnieje około 300 000 gatunków roślin kwiatowych. Stanowią dwie trzecie wszystkich istniejących dziś gatunków roślin i dominują w większości siedlisk na lądzie. Cieszymy się częścią tej różnorodności, gdy spacerujemy z otwartymi oczami po kwitnącej łące lub po pięknie zagospodarowanym ogrodzie. [MM]

Kolorowa kwietna łąka, sfotografowana w Heidelbergu (źródło:
https://commons.wikimedia.org/wiki/File:HD_Bunte_Blumenwiese_2011.JPG)
{kind=link}