







130 Millionen Jahre vor heute
![]()
Zeitalter: Mesozoikum / Kreide
 Die Entwicklung der Blütenpflanzen hat vermutlich bereits vor ca. 200 Mio. Jahren begonnen, indem es in einigen Pflanzen zu einer Vervielfachung ihres gesamten Erbmaterials kam. Weitere Veränderungen der vervielfachten Chromosomen führten zur Ausbildung von Blüten. Diese Entwicklung muss mehrmals stattgefunden haben, was die Vielzahl der grundlegenden Blütenbaupläne zeigt.
Die Entwicklung der Blütenpflanzen hat vermutlich bereits vor ca. 200 Mio. Jahren begonnen, indem es in einigen Pflanzen zu einer Vervielfachung ihres gesamten Erbmaterials kam. Weitere Veränderungen der vervielfachten Chromosomen führten zur Ausbildung von Blüten. Diese Entwicklung muss mehrmals stattgefunden haben, was die Vielzahl der grundlegenden Blütenbaupläne zeigt.
Die ältesten versteinerten Blütenpflanzen sind allerdings nur etwa 130 Mio. Jahre alt und sehr spärlich. Daher ist ihre Entwicklung schlecht erforscht. Möglicherweise haben sie sich aus Farnen entwickelt, die schon seit 400 Mio. Jahren nachweisbar sind. Die Blütenpflanzen haben bis heute Einfluss auf die Entwicklung der Insekten und umgekehrt.
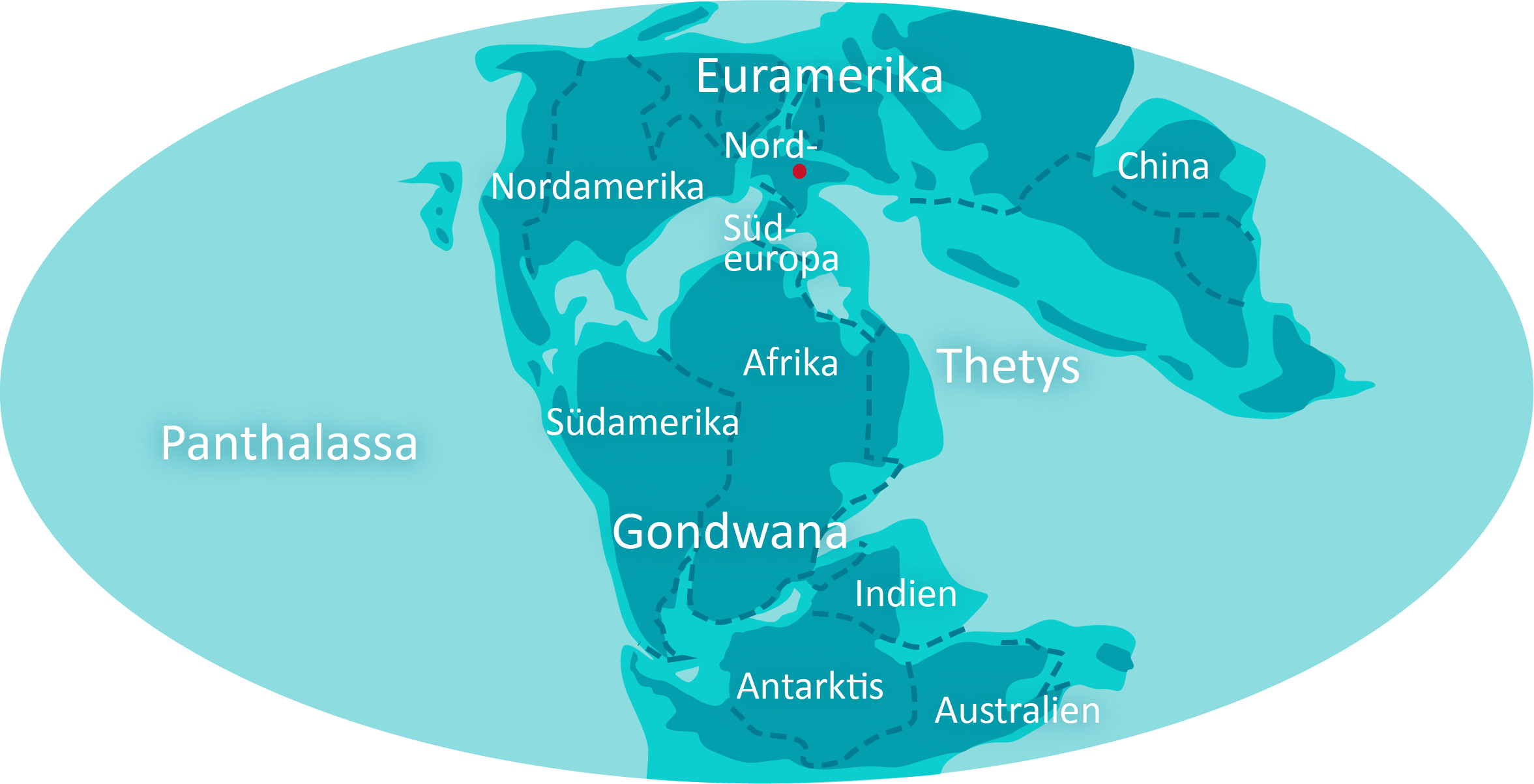 Das Klima ist durchgehend warm. Das Land ist eisfrei, deshalb ist der Meeresspiegel hoch. Die Atmosphäre enthält 1,5‑mal soviel Sauerstoff und 4‑mal soviel CO2 wie heute. Der Zerfall von Gondwana setzt sich fort. Australien trennt sich von Antarktika und Afrika von Südamerika, auch Indien spaltet sich ab.
Das Klima ist durchgehend warm. Das Land ist eisfrei, deshalb ist der Meeresspiegel hoch. Die Atmosphäre enthält 1,5‑mal soviel Sauerstoff und 4‑mal soviel CO2 wie heute. Der Zerfall von Gondwana setzt sich fort. Australien trennt sich von Antarktika und Afrika von Südamerika, auch Indien spaltet sich ab.
Diese Infos in einfacher Sprache auf evokids.de.
Diese Station im Hörspiel:
Die Gruppe der Samenpflanzen umfasst praktisch alle heute lebenden Landpflanzen mit Ausnahme der Farne und Moose. Die ersten Samenpflanzen sind bereits im Devon entstanden. Im Karbon haben sie Stützskelette entwickelt, so dass sie größere Wuchsformen ausbilden konnten.
Als „Blütenpflanzen“ wird in aller Regel die Gruppe der „Bedecktsamer“ (Angiospermen) bezeichnet. Ihre genaue Abstammung ist bis heute ungeklärt. Man geht davon aus, dass sie sich, ebenso wie die Gruppe der „Nacktsamer“ (Gymnospermen), aus der gemeinsamen Stammgruppe der Samenfarne (auch Farnsamer genannt) entwickelt haben. Wie der Name sagt, ist ihnen allen die Fähigkeit zur Samenbildung gemeinsam.
Älteste Fossilien (Pollen und Blattreste) weisen auf eine Entstehung erster Bedecktsamer bereits im Jura hin. Eine geradezu explosive Evolution der Blütenpflanzen mit einer hohen Artbildungsrate fand jedoch in der Kreidezeit statt.
Dominanz der Nacktsamer
Bis zum Ende des Jura beherrschten, neben Moosen und Farnen sowie verschiedenen ursprünglichen Pflanzenformen, zu denen auch die heute noch lebenden Palmfarne zählen, die bereits erwähnten „Nacktsamer“ (Gymnospermen) die Landschaft. Zu ihnen gehören, neben dem Ginkgo als „lebendem Fossil“, beispielsweise alle heute noch lebenden Koniferen, die Zypressengewächse und Wacholder-Arten. Ihre Samenanlagen liegen offen auf den Fruchtblättern (Samenschuppen).
Diese wiederum sind häufig zu kreisförmigen, eng übereinandergelagerten Blütenständen an Kurzsprossen zusammengeschlossen. Die Kurzsprosse und Samenschuppen der weiblichen Blüten sind übrigens häufig verholzt, was am Beispiel der Kieferzapfen gut zu erkennen ist.
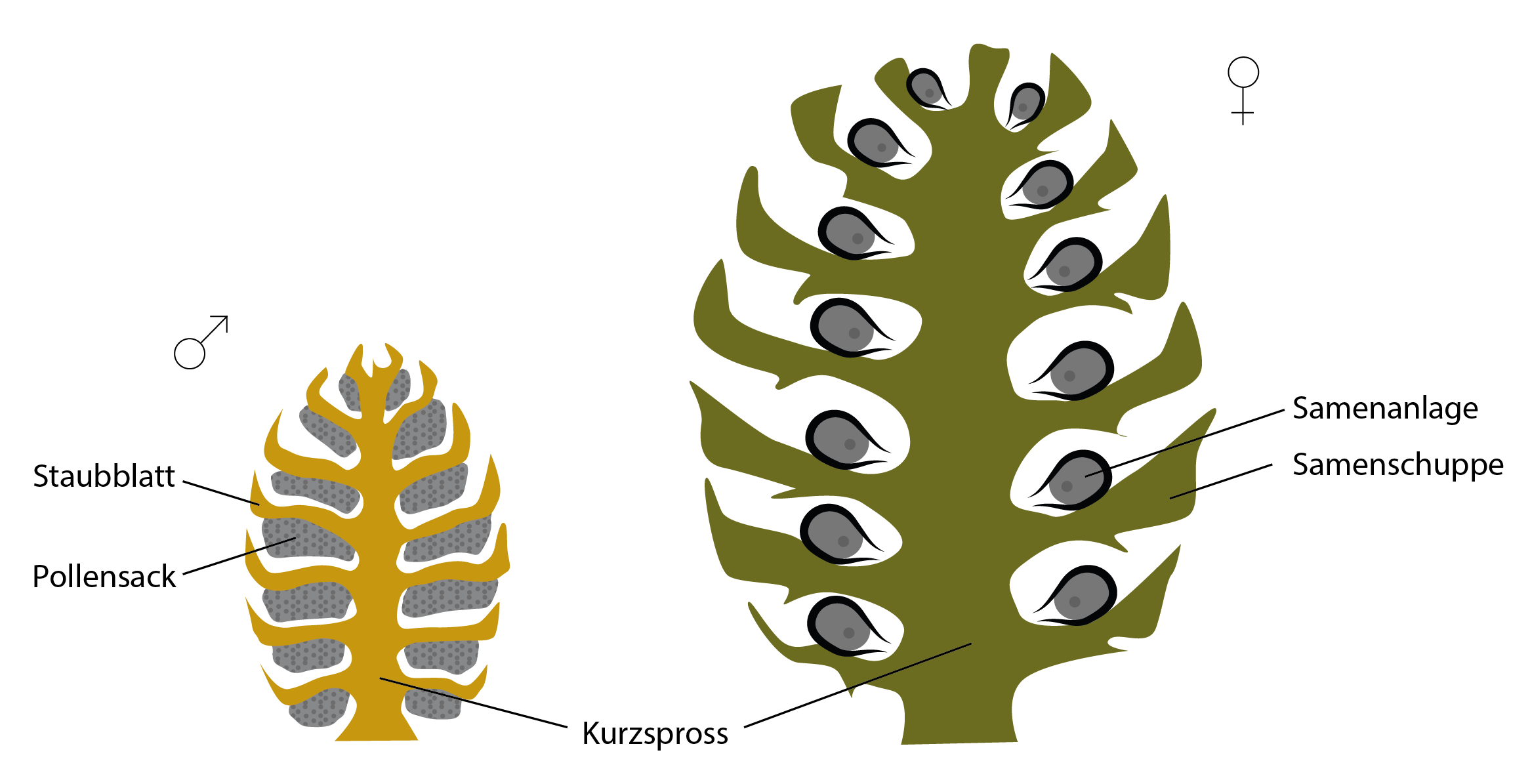
Nacktsamer-Blütenstand (links männlich, rechts weiblich). Die Samenanlagen des weiblichen Blütenstandes liegen unbedeckt auf der Samenschuppe.

Nacktsamer-Blüten. Links: blühender Lärchenzapfen; rechts: Fichtenzapfen. (Quellen: https://commons.wikimedia.org/wiki/File:Kywiat.jpg, http://commons.wikimedia.org/wiki/File:Picea_Abies_female_cone_cropped.jpg)
{kind=link}
{kind=link}
Die Befruchtung der weiblichen Nacktsamer-Blüten erfolgt in aller Regel durch Windbestäubung, gelegentlich sind auch Insekten (meist Käfer) beteiligt.
Vervielfachte Chromosomen – Spielraum für Neuentwicklungen
Bereits vor etwa 320 Millionen Jahren kam es in Vorläufern der Samenpflanzen zu einer Vervielfachung des gesamten Erbmaterials. Vor etwa 200 Millionen Jahren ereignete sich eine solche Vervielfachung auch bei den Vorfahren der heutigen Blütenpflanzen. Durch umfangreiche Vergleiche von Gengruppen in heutigen Blütenpflanzen, Moosen und Moosfarnen, die aus vervielfachten Genen hervorgingen, hat man die Zeiträume dieser Vervielfachungen eingekreist.
Während tierische Organismen mit mehrfach vorhandenen Chromosomen (mit wenigen Ausnahmen einzelner weniger Trisomien) in der Regel nicht lebensfähig sind, scheint dies Planzenzellen keine Probleme zu bereiten. Sie profitieren sogar von den vervielfachten Chromosomen: Viele der mehrfach vorhandenen Gene verschwanden zwar mit der Zeit, andere veränderten sich jedoch und übernahmen neue Funktionen. Unter anderem steuerten sie neu entstandene Prozesse rund ums Blühen.
Aufkommen der Bedecktsamer
In der Kreidezeit haben sich Pflanzen entwickelt, die ihre Samenanlagen nicht mehr offen tragen, sondern deren Fruchtblatt eingerollt ist. Ein Vorteil dieser Entwicklung war, dass die Samenanlagen geschützt im Inneren liegen. Pflanzen, bei denen dies der Fall ist, bezeichnet man als „Bedecktsamer“ (Angiospermen) oder einfach als „Blütenpflanzen“. Bei Pflanzen mit mehreren Fruchblättern pro Blüte sind diese auf vielfältige Weise miteinander verwachsen – auch hier immer mit dem Ergebnis, dass die Samenanlagen geschützt im Inneren zu liegen kommen. So können Tiere als Bestäuber dienen, ohne dass die Samenanlage gleichzeitig stark fraßgefährdet ist.
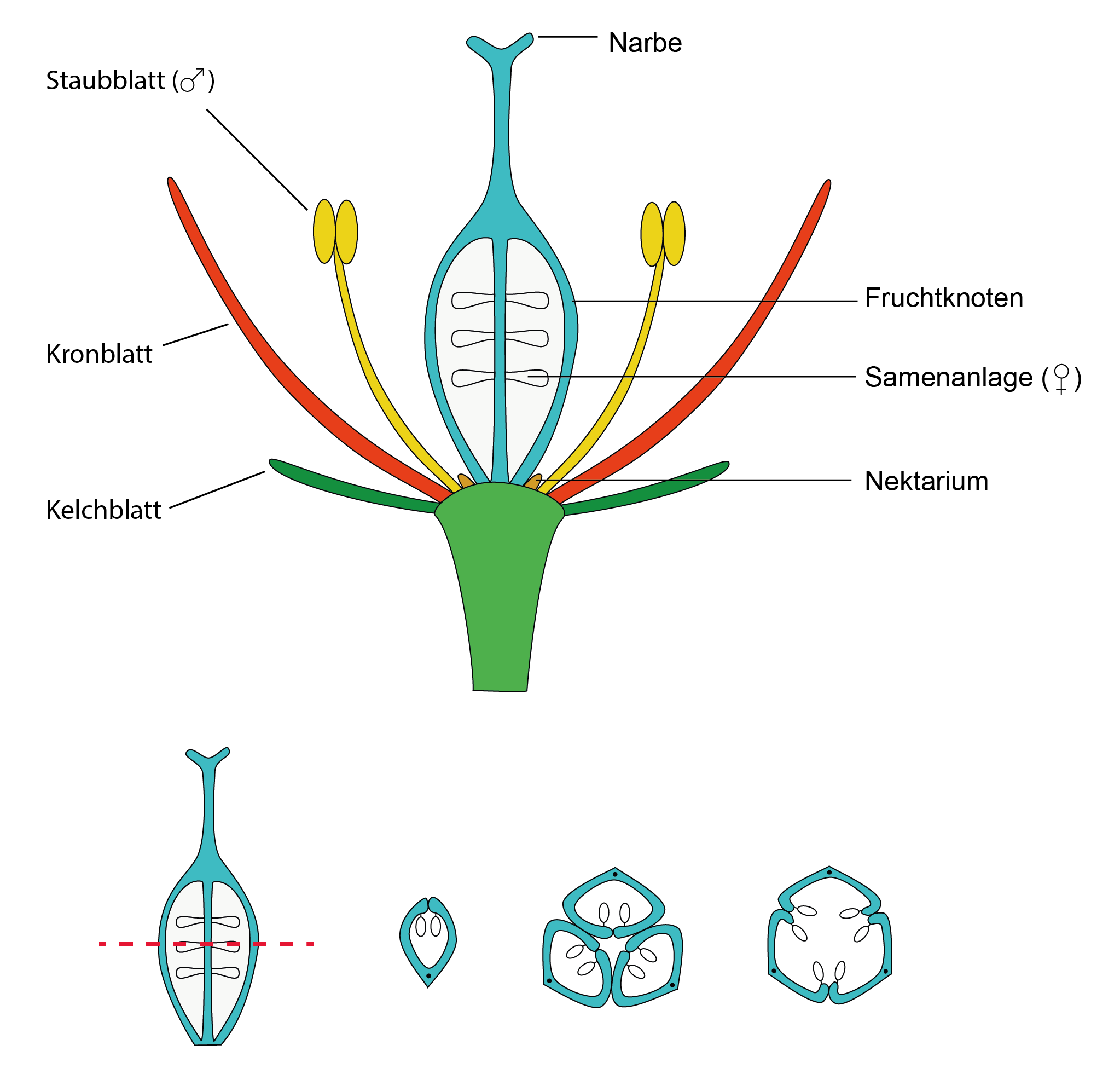
Bauplan einer Bedecktsamer-Blüte. Die Blüte wird aus spezialisierten Blättern gebildet. Die Kronblätter sind häufig sehr vielgestaltig. Die Staubblätter produzieren den (männlichen) Pollen, die Fruchtblätter tragen die (weiblichen) Samenanlagen und sind zum Fruchtknoten verwachsen. Im unteren Bildteil sind drei von vielen Möglichkeiten des Fruchtknotenaufbaus gezeigt.
Der Pollen trifft nicht mehr unmittelbar auf die Samenanlage, sondern auf den als Narbe bezeichneten Teil der verwachsenen Fruchtblätter. Dort bildet er einen Schlauch aus, in dem die beiden männlichen Samenzellen des Pollens zu der weiblichen Samenzelle und ihren Begleiterinnen im Inneren der Samenanlage gelangen.
Dieses Konstrukt bringt mit sich, dass nur jener Pollen von den vielen, die auf der Narbe landen, die Befruchtung vollziehen kann, der tatsächlich in der Lage ist, am schnellsten einen stabilen Pollenschlauch auszubilden.
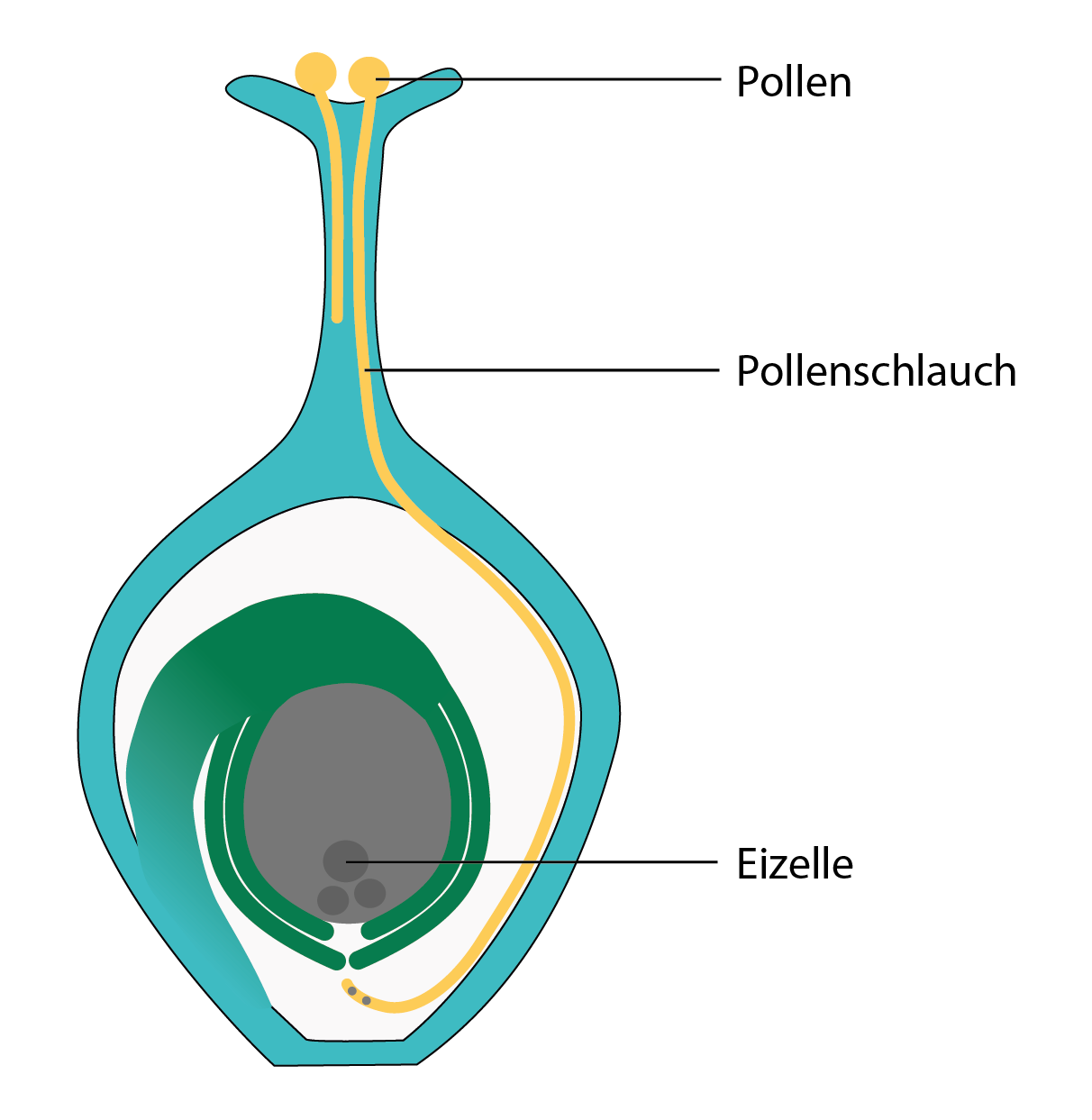
Pollenschlauch. Der Pollen bildet einen Schlauch, in dem seine Kerne zur Eizelle transportiert werden. Die weibliche Samenanlage liegt, geschützt von Hüllstrukturen, im inneren des Fruchtknotens.
Nicht nur im Vorhandensein der Blütenorgane und ihrem Aufbau unterscheiden sich die Bedecktsamer von den anderen Pflanzengruppen. Auch viele weitere Merkmale tragen zur Differenzierung bei, wie etwa die Pollenmorphologie, eine charakteristische Differenzierung des Leitgewebes sowie eine Vielfalt sekundärer Pflanzenstoffe, z.B. Duftstoffe oder Fraßabwehrstoffe.
Nährgewebe für den Embryo: Ökonomik der doppelten Befruchtung
Charakteristisch ist auch, dass im Zuge der Befruchtung bei Bedecktsamern beide Spermakerne aus dem Pollenschlauch in die Samenanlage gelangen (sog. doppelte Befruchtung), von denen der eine die weibliche Eizelle befruchtet, der andere dem bereits mit zwei Kernen versehenen sog. Embryosack einen dritten Kern hinzufügt. Dieser dann triploide Embryosack teilt sich und entwickelt sich zu einem Nährgewebe für den Embryo (sog. sekundäres Endosperm).
Bei den Gymnospermen findet hingegen die Einlagerung von Speicherstoffen in die Samenanlage häufig bereits vor der Befruchtung statt (z.B. Pinienkerne, Cedernüsse). Dieses haploide Nährgewebe bezeichnet man als primäres Endosperm.
Der Vorteil, wenn erst nach der Befruchtung Energie in den Aufbau des Nährgewebes investiert wird, liegt auf der Hand: Neben der Tatsache, dass das Nährgewebe der Nacktsamer stark fraßgefährdet ist, ist es zudem vergeudet, wenn es nicht zur Befruchtung kommt. Wird dagegen seine Bildung erst gemeinsam mit der Befruchtung angestoßen, geschieht dies genau dann, wenn es tatsächlich benötigt wird.
Coevolution von Blüten, Bestäubern und Samenverteilern
Ergänzend zur Entwicklung eines schützenden Fruchtknotens haben sich bei Angiospermen-Blüten vielfältige Blütenblatt-Spezialisierungen herausgebildet, die bestäubende Insekten anlocken. Weitere Errungenschaften, wie Nektar- und Duftdrüsen, tragen ihr Übriges dazu bei.
Dabei haben sich sehr spezielle Wechselbeziehungen zwischen Insekten und Pflanzen herausgebildet. So sind manche Insekten auf eine ganz bestimmte Pflanzenart fokussiert, durch die sie angelockt werden, und haben ihrerseits entsprechende Organe entwickelt (z.B. lange Saugrüssel bei Schmetterlingen oder Pollensäcke bei Bienen), um an die Lockstoffe der Pflanzen zu gelangen und bei dieser Gelegenheit – als Nutzen für die Pflanze – häufig die Befruchtung sicherzustellen.
Mehr Informationen und vor allem schöne Bilder dazu finden sich z.B. hier http://deinnachbarlohbach.blogspot.com/2015/04/blutenbestauber.html
Auch zur Verbreitung der befruchteten Samen haben sich zwischen Pflanzen und Tieren im Laufe der Evolution faszinierende Kooperationen herausgebildet. Um eine möglichst weite Verbreitung zu erreichen, bilden viele Pflanzen aus Teilen ihrer Blüte fleischige Früchte um ihre Samen herum. Diese werden von Tieren gefressen und die Samen an entfernter Stelle wieder ausgeschieden. Bei einigen Samen erhöht die Passage durch den Verdauungstrakt des verbreitenden Tieres sogar die Wahrscheinlichkeit, dass sie überhaupt erfolgreich auskeimen.
Auch der Mensch macht sich die Vielfalt der Angiospermenfrüchte zunutze. So sind Obst, Gemüse, Getreide wichtige Bestandteile unserer Ernährung, und auch viele andere Teile dieser Pflanzen (Fasern, Sekundärstoffe) sind für uns als Rohstoffe für Kleidung, Papier oder Arzneien unentbehrlich.
Die beiden großen Gruppen der Blütenpflanzen
Die Angiospermen werden traditionell in zwei große Gruppen eingeteilt, basierend auf der Zahl ihrer Keimblätter:
- Die sogenannten Einkeimblättrigen bilden nach der Keimung ein einzelnes Keimblatt. Ihre Blätter sind in der Regel länglich, die Blattadern parallel angeordnet, und die Blüten haben eine 3er-Symmetrie (jeweils 3 Kelchblätter, Kronblätter, Staub- und Fruchtblätter oder Vielfache davon). Beispiele sind alle Gräser, aber auch Lilien und Tulpen.
- Die sogenannten Zweikeimblättrigen bilden, wie der Name sagt, zwei Keimblätter. Ihre Blattadern sind verzweigt, und die Blüten haben häufig eine 4er- (z.B. Raps) oder 5er-Symmetrie (z.B. alle Obstgehölze).
Weltweit gibt es heute etwa 300000 Arten von Blütenpflanzen. Damit stellen sie zwei Drittel aller heute existierenden Pflanzenarten und dominieren die meisten Lebensräume auf dem Land. An einem Teil dieser Vielfalt erfreuen wir uns, wann immer wir offenen Auges über eine blühende Wiese oder durch einen schön angelegten Garten gehen. [MM]

Bunte Blumenwiese, fotografiert in Heidelberg (Quelle:
https://commons.wikimedia.org/wiki/File:HD_Bunte_Blumenwiese_2011.JPG)
{kind=link}