







450 milionów lat przed dniem dzisiejszym
![]()
Epoka: paleozoik / ordowik
 Pierwsze rośliny i owady przemieszczają się na ląd, pojawia się wiele nowych gatunków. Klimat jest tropikalny, ale rośliny zmieniają go zasadniczo: wiążą dwutlenek węgla (gaz cieplarniany) z atmosfery, i temperatura na ziemi spada. Jednocześnie wzrasta zawartość tlenu w atmosferze. Zmienia się również chemia morza. Główny kontynent Gondwany dryfuje nad biegunem południowym. Wszystko to razem powoduje drugie wielkie wymieranie gatunków w historii Ziemi.
Pierwsze rośliny i owady przemieszczają się na ląd, pojawia się wiele nowych gatunków. Klimat jest tropikalny, ale rośliny zmieniają go zasadniczo: wiążą dwutlenek węgla (gaz cieplarniany) z atmosfery, i temperatura na ziemi spada. Jednocześnie wzrasta zawartość tlenu w atmosferze. Zmienia się również chemia morza. Główny kontynent Gondwany dryfuje nad biegunem południowym. Wszystko to razem powoduje drugie wielkie wymieranie gatunków w historii Ziemi.

Superkontynent Gondwana tworzy dużą masę lądową na półkuli południowej. Dalej na północ zbliżają się do siebie Laurencja i Bałtyka. Duża część kontynentów jest początkowo pokryta płytkimi, ciepłymi morzami. Jednak spadek temperatury spowodowany metabolizmem roślin prowadzi do nowej epoki lodowcowej.
Tę informację w prostym języku znajdziesz na stronie evokids.de.
Czas, w którym rośliny wyszły na ląd, jest dobrze udokumentowany przez liczne znaleziska skamieniałości. Prawdopodobnie miało to miejsce we ordowiku, około 480–450 milionów lat temu, a więc na dobre 100 milionów lat przed wyjściem na ląd kręgowców. Wyjście roślin na ląd prawdopodobnie miało bezpośredni i znaczący wpływ na rozwój klimatu, ponieważ rosnąca aktywność fotosyntetyczna zmniejszyła stężenie CO2 w atmosferze, a to doprowadziło do ochłodzenia klimatu tropikalnego. Jednak potwierdzone znaleziska skamieniałości są ograniczone do gatunków, które były już dobrze przystosowane do życia na lądzie. Określenie wieku metodą zegara molekularnego sugeruje nawet, że najstarsze rośliny lądowe były o około 300 milionów lat starsze od najstarszych skamieniałych okazów.
Jest bardzo prawdopodobne, że pierwsze rośliny lądowe rozwinęły się z grupy ramienic właściwych (Charales), które mają podobieństwa we wzroście, rozmnażaniu i podziale komórek z roślinami lądowymi. Potwierdziły to molekularne badania genetyczne, w których porównano genomy czterdziestu różnych glonów i roślin lądowych.

Ramienica pospolita (Chara vulgaris) tworzy dywan w pobliżu powierzchni wody w stawie. (Źródło: https://commons.wikimedia.org/wiki/File:Chara1.jpg)
Wymagania do życia na lądzie
Chociaż rośliny wodne nigdy nie doświadczają stresu związanego z suszą, jest to stałe zagrożenie dla roślin lądowych. Mechanizm rozwinięty w tym celu przez rośliny lądowe polega na wytwarzaniu hormonu stresu, kwasu abscysynowego. Jest on uwalniany w dzisiejszych roślinach w odpowiedzi na suszę i zapewnia przejście w tryb oszczędzania wody. Taka regulacja jest zbędna w przypadku ramiennic żyjących w wodzie, ale mają one już geny kodujące wczesne etapy syntezy fitohormonu. Ramiennice są również w stanie wykryć etylen, który we współczesnych roślinach kontroluje takie procesy jak rozwój kwiatów czy dojrzewanie owoców; Jednak (jeszcze) brakuje im genów do produkcji tego gazu.
Tworzenie korzenia i oskórka (kutykula)
Dzisiejsze rośliny lądowe mają zwykle oskórek, woskową warstwę pokrywającą całą roślinę (z wyjątkiem korzeni) i oddzieloną od zewnętrznej warstwy komórek.
Pierwsze rośliny lądowe nie miały wcale (lub przynajmniej sprawnego) oskórka i mogły wchłaniać wodę i składniki odżywcze całą powierzchnią. Nie miały też jeszcze korzeni. Ich ryzoidy, czasami postrzegane jako analogony korzeni, nie powstały głównie do wchłaniania wody i składników odżywczych, ale były używane do przyczepiania się do podłoża. Roślinami żyjącymi dzisiaj z takimi ryzoidami, czyli bez prawdziwych korzeni, są mchy.
Pierwsze przystosowania do życia na wsi polegały na tym, że fragmenty roślin wyniesionych nad ziemię były chronione przed utratą wody poprzez tworzenie się oskórka. W tym samym czasie absorpcja wody została przesunięta na niższe obszary roślin.

Schemat budowy rośliny mchu. Składniki odżywcze i woda są wchłaniane przez dolne liście. Ryzoid służy jedynie do zakotwiczenia go w ziemi.
Dopiero u paproci pojawiają się przerosty tkanek we wnętrzu rośliny, co prowadzi do pękania naskórka i powoduje pęcznienie tkanki. Tkanka ta nie może być od samego początku pokryta ochronną warstwą kutiny, dlatego idealnie nadaje się do wchłaniania wody i składników odżywczych. Dzięki ryzoidom, które utrzymują roślinę na podłożu, tkanka wyrastająca z wnętrza organizmu jest wciskana w podłoże i nie unosi rośliny. Jednak te wyrostki komórkowe wczesnych paproci nie są jeszcze prawdziwymi korzeniami. Na przykład nie mają wewnątrz przewodzącej tkanki, w której mógłby odbywać się ukierunkowany transport wody lub substancji.

Paproć pospolita z kłączem i liśćmi. Małe sekcje po prawej stronie pokazują pojemniki z zarodnikami, które rosną w punktowych skupiskach na spodniej stronie liści. (Źródło: https://de.wikipedia.org/wiki/Echte_Farne#/media/File:Dryopteris_filix_mas_nf.jpg)
Powstawanie prawdziwych korzeni nie jest dobrze udokumentowane przez skamieniałości. Obecnie istnieją dwa zasadniczo różne systemy korzeniowe, a mianowicie jednorodne systemy korzeniowe paproci i roślin jednoliściennych, w których wiele podobnych nitek korzeni pojawia się na dolnym końcu osi łodygi oraz heterogeniczne systemy korzeni dwuliściennych, w których jeden lub więcej głównych korzeni rozgałęzia się w korzenie boczne. Jednorodne systemy korzeniowe paproci były prawdopodobnie pierwszymi prawdziwymi korzeniami, w których ukształtowały się tkanki przewodzące do transportu wody i składników odżywczych do organizmu rośliny.
Podczas kiełkowania korzenie najpierw wyrastają z okrywy nasiennej i zapewniają zakotwiczenie sadzonki w ziemi oraz dostarczanie roślinom składników odżywczych. W przypadku korzenia wnikanie w podłoże, takie jak gleba, jest również ułatwione przez powstawanie śliskiej warstwy z degenerujących się (ginących) komórek, które nieustannie wykształcają się na wierzchołku korzenia. Taka czapka korzeniowa lub kaliptra nad wierzchołkową strefą wzrostu korzenia jest zawsze obecna, nawet przy korzeniach powietrznych, jako ochrona dla komórek wyrastających z tkanki.
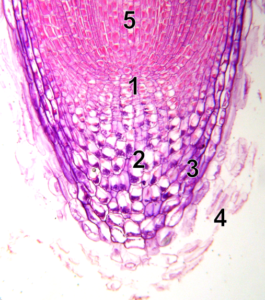
Korzeń prawdziwy z kopułą z martwych komórek (4) (źródło: https://commons.wikimedia.org/wiki/File:Root-tip-tag.png)
Adaptacja reprodukcji
Aby zabezpieczyć kiełkowanie nowej rośliny przed wysychaniem, rośliny lądowe umieściły pierwsze podziały komórkowe tych zarodków w osłonie (płaszczu nasiennym). Ta otoczka została utworzona jeszcze przez roślinę matkę i zawiera również tkankę odżywczą dla zarodka, który w niej rośnie. Zarodek w skorupie nazywany jest również embrionem, dlatego rośliny lądowe nazywane są embriofitami.

Embrion roślinny. Zarodek rośnie z zapłodnionej zygoty do tkanki odżywczej (bielma), która również się rozmnaża. W okrywie nasiennej powstają liścienie i korzeń (radicula). Przykład przedstawia roślinę dwuliścienną.
Tworzenie embrionów u roślin i zwierząt rozwijało się niezależnie od siebie i powinno być postrzegane jako metoda rozwoju zarodków poza wodą. [KHB]