







560 milionów lat przed dniem dzisiejszym
![]()
Epoka: neoproterozoik / ediakar
 Z tego okresu pochodzą pierwsze skamieniałości skupisk połączonych komórek. Dzięki łączeniu się tworzą podstawę do podziału pracy między komórkami i rozwoju bardziej złożonych żywych istot.
Z tego okresu pochodzą pierwsze skamieniałości skupisk połączonych komórek. Dzięki łączeniu się tworzą podstawę do podziału pracy między komórkami i rozwoju bardziej złożonych żywych istot.
Białka strukturalne zapewniają szybko rozwijającym się istotom żywym formę i wsparcie. Wbudowanie wapna w szkielety tworzy solidną zbroję, która nie tylko wspiera organizmy, ale także zapewnia ochronę przed urazami. Dzięki temu mogą powstawać większe formy życia. Powstaje wiele podstawowych planów żywych istot, które dziś nadal istnieją.
Na pierwszym planie ilustracji widać trylobita Cheirurus ingricus.
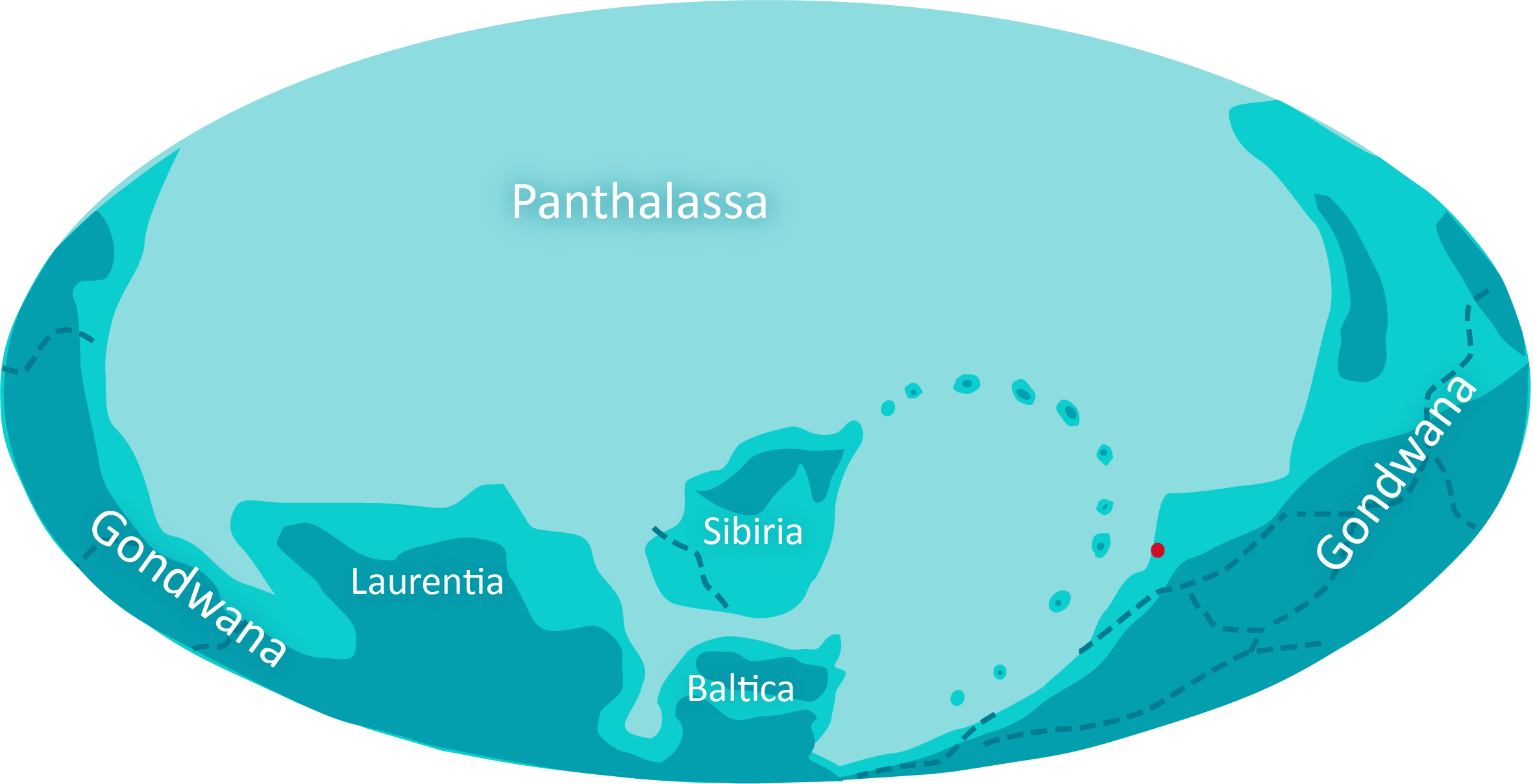
Przemieszczanie się mas lądowych w pobliżu bieguna przerywa cykl przemiennego oblodzenia i przegrzania ziemi. Zawartość CO2 w atmosferze stabilizuje się, a wraz z nią temperatura.
Tę informację w prostym języku znajdziesz na stronie evokids.de.
Obecnie szkielety wspierające i ochronne obejmują egzoszkielet, endoszkielet i hydroszkielet.
Najstarszym skamieniałym zwierzęciem z podtrzymywanym szkieletem jest Namapoikia reitogensis. Nie wiadomo, czy był to koralowiec, czy gąbka, jego wiek szacuje się na około 550 milionów lat.
Kiedy pierwsze większe organizmy wielokomórkowe pojawiły się około 560 milionów lat temu, mogło to nastąpić tylko jednocześnie z rozwojem i powstawaniem szkieletów wspierających i ochronnych, co miało miejsce zarówno w królestwie zwierząt, jak i roślin. Podczas gdy lignina rozwinęła się jako najważniejsza cząsteczka podtrzymująca w roślinach i została wbudowana obok złogów mineralnych między włóknami celulozowymi ścian zewnętrznych, u wielu zwierząt chityna rozwinęła się w celu zewnętrznej stabilizacji organizmu. Podobnie jak celuloza, chityna jest polisacharydem, ale składa się z monomerów N‑acetyloglukozaminy, która składa się również z glukozy, ale nadal zawiera resztę acetylowanej aminy.
Chityna prawdopodobnie pochodzi z bakterii, w których jest głównym składnikiem ściany komórkowej.
Dlaczego komórki rosną (kiedy mogą)?
Komórki stają się większe, ponieważ jest to w wielu przypadkach korzystne termodynamicznie, tj. fizycznie i energetycznie. Stosunek objętości do powierzchni komórki stanowi jedną trzecią promienia kuli:

Oznacza to, że im większa staje się kula, tym bardziej rośnie jej zawartość w porównaniu z jej powierzchnią. Oznacza to, że wymiana wody, materii i ciepła z otoczeniem może być lepiej kontrolowana. To zjawisko wzrostu wielkości można zaobserwować u wielu organizmów jednokomórkowych, które żyją przy ograniczonej podaży składników odżywczych. Jednak zwiększenie objętości ma również wady, np. siły, które występują przy różnicach ciśnień osmotycznych między wnętrzem i zewnętrzem komórki są znacznie większe. W tym przypadku może pomóc ochrona mechaniczna. Komórki bez takiej ochrony mogą istnieć tylko w równowadze osmotycznej z otoczeniem i są również wrażliwe na mechaniczne siły ścinające.
Już jednokomórkowce, zarówno prokariotycznejak i eukariotyczne, rozwinęły ścianę komórkową zbudowaną z białek pomocniczych i polisacharydów, tak zwanych peptydoglikanów, które zapewniają ich komórkom mechaniczną ochronę na wczesnym etapie ewolucji. Stało się to co najmniej 3,5 miliarda lat temu, jak pokazują znaleziska skamieniałości prekursorów cyjanoficea. Jedynymi wyjątkami są bakterie pasożytnicze, takie jak mykoplazmy i tym podobne.
Jednak taka ściana komórkowa nie jest już wystarczająca dla większych grup komórek i organizmów wielokomórkowych. Dopiero utworzenie dalszych struktur stabilizujących rozwiązało ten problem.
Egzoszkielet (szkielet zewnętrzny)
Najwcześniejszą z tych struktur była celuloza, która składa się z długich łańcuchów czystych cząsteczek glukozy (cukru gronowego) połączonych specjalnym wiązaniem chemicznym. Celuloza ta powstaje w błonie komórkowej komórek, osadza się na zewnątrz i jej cząsteczki natychmiast łączą sie. Z jednym wyjątkiem, tunicata, tylko komórki roślinne są zdolne do syntezy i wydzielania (wydalania) celulozy. Dzięki swojej strukturze włókno celulozowe może wytrzymać duże obciążenia rozciągające, ale nie wytrzymuje prawie żadnego nacisku (rośliny osiągnęły stabilność ściskanie tylko dzięki magazynowaniu ligniny między włóknami celulozowymi).
W świecie zwierząt, z wyjątkiem wspomnianych wyjątków, egzoszkielet powstawał zwłaszcza u stawonogów jako zewnętrzne złogi naskórka. Nazywa się naskórek i jako istotny element strukturalny składa się z bezbarwnych, elastycznych włókien chitynowych. Składnik białkowy, brązowawa sklerotyna, otacza te fibryle i określa wytrzymałość kompleksu. Minerały, takie jak kalcyt lub apatyt, mogą być również magazynowane jako dodatkowe wzmocnienie, ale także mogą to być związki cynku lub magnezu. Mięśnie przyczepiają się od wewnątrz do poszczególnych elementów egzoszkieletu, umożliwiając ruch elementów względem siebie.
Ponieważ egzoszkielety są nierozbieralne i nie rosną razem z nimi, stawonogi są zmuszone opuścić tę skorupę w fazie wzrostu i uformować nowy, większy egzoszkielet. Ten proces nazywa się „linieniem”. Ponieważ pancerz chitynowy nie chroni przed utratą wody, u wielu stawonogów na naskórku odkłada się woskowata warstwa ochronna zwana Epicuticula.
Hydroszkielet (szkielet hydrostatyczny)
Hydroszkielet jest filogenetycznie starszy niż egzoszkielet, tj. powstał przed nim w dziejach ewolucji. Mówi się o hydroszkielecie, gdy część mięśni działa na doszczętnie wypełnioną poduszkę z płynem, dzięki czemu siły mechaniczne mogą być przyjmowane i skutecznie dalej przekazywane, podobnie jak w technicznym układzie hydraulicznym. Ten system prawdopodobnie wywodzi się z nicieni i pierścienic i istnieje do dziś.
Owady również wykorzystują ten hydroszkielet, pomimo swojego uksztatowanego egzoszkieletu, np. do poruszania nóg do skakania lub do chodzenia, a także do pozbycia się ich egzoszkieletu podczas procesów linienia. Funkcjonalnie hydroszkieletowi odpowiada tzw. turgor komórkowy roślin, w którym ciśnienie osmotyczne wnętrza komórki, zwiększone w porównaniu z ośrodkiem zewnętrznym, prowadzi do hydrostatycznej różnicy ciśnień, która musi zostać przejęta przez ściankę komórkową.
Endoszkielet (szkielet wewnętrzny)
Endoszkielet czyli szkielet wewnętrzny jest najbardziej zauważalną wspólną cechą kręgowców. Jest to mechaniczna konstrukcja nośna (szkielet), która nie jest częścią zewnętrznej powłoki organizmu. Jego poszczególne elementy nazywane są kośćmi. Podczas gdy u stawonogów mięśnie przyczepione są do wnętrza egzoszkieletu, u kręgowców przyczepiają się do kości wewnątrz ciała i poruszają poszczególne części ciała względem siebie. Z wyjątkiem ryb chrzęstnych, których szkielet wewnętrzny składa się z chrząstki z włączonym węglanem wapnia, szkielet wewnętrzny większości kręgowców składa się z fosforanu wapnia i kolagenu. Kolejnym składnikiem wewnętrznego szkieletu jest chrząstka. W przeciwieństwie do egzoszkieletu zbudowanego z chityny, taki kościsty szkielet wewnętrzny rośnie wraz z organizmem: każda chrząstka i każda kość podlega ciągłemu rozpadowi i odbudowie. [KHB]