







2500 milionów lat przed dniem dzisiejszym
![]()
Epoka: proterozoik / sider
 Komórki rozwijają zdolność coraz wydajniejniejszego wytwarzania energii ze światła słonecznego. Powstający tlen przez miliony lat prowadzi jedynie do tworzenia nierozpuszczalnych w wodzie soli metali w oceanach. Powstają potężne warstwy rudy odkrywkowej, które nadal eksploatujemy.
Komórki rozwijają zdolność coraz wydajniejniejszego wytwarzania energii ze światła słonecznego. Powstający tlen przez miliony lat prowadzi jedynie do tworzenia nierozpuszczalnych w wodzie soli metali w oceanach. Powstają potężne warstwy rudy odkrywkowej, które nadal eksploatujemy.
Stężenie żelaza w wodzie spada, tak że występuje wolny tlen. Jest on jednak trujący: pierwsze poważne wymieranie („wielka katastrofa tlenowa”) dotyka większość gatunków. Organizmy żyjące w środowisku beztlenowym pozostają nienaruszone. Rozprzestrzeniają się nowe gatunki, które tolerują tlen lub mogą nawet czerpać z niego dużo energii.
 Pierwszy tlen dostaje się do atmosfery, jego udział stale rośnie. Jednocześnie spada zawartość dwutlenku węgla. Ziemię w większości pokrywają oceany. Tylko kilka stożków wulkanicznych lub wzniesionych fragmentów skorupy ziemi (kratonów) góruje nad powierzchnią wody.
Pierwszy tlen dostaje się do atmosfery, jego udział stale rośnie. Jednocześnie spada zawartość dwutlenku węgla. Ziemię w większości pokrywają oceany. Tylko kilka stożków wulkanicznych lub wzniesionych fragmentów skorupy ziemi (kratonów) góruje nad powierzchnią wody.
Tę informację w prostym języku znajdziesz na stronie evokids.de.
Fotosynteza jest najważniejszą reakcją dla życia na Ziemi w jego obecnej formie. Jej znaczenia nie można przecenić, ponieważ umożliwia ona wszystkim stworzeniom zwierzęcym oraz wielu bakteriom i grzybom, a także oczywiście samym organizmom prowadzącym fotosyntezę, wykorzystanie energii słonecznej jako podstawy do życia. Zdecydowana większość organizmów żyjących obecnie jest zależna od produktów fotosyntezy służących im do napędu własnego metabolizmu.
Obecnie fotosynteza zachodzi na niezwykle wielką skalę i prowadzona jest głównie przez lądowe rośliny zielone. Wiążą one co roku około 120 miliardów ton CO2, z czego około połowa jest związana jest w postaci biomasy (druga połowa związanego CO2 jest ponownie uwalniana w trakcie metabolizmu roślin i uwalniana do atmosfery). Ponadto każdego roku około 55 miliardów ton biomasy jest fotosyntetyzowanych przez organizmy morskie.
2,5 miliarda lat temu oczywiście nie było jeszczewyższych roślin, nie rozwinęły się nawet jednokomórkowce z jądrem komórki. W tym wczesnym okresie życia fotosynteza tlenowa (uwalniająca tlen) została „wynaleziona” przez cyjanobakterie, które wraz z osadami bogatymi w minerały tworzyły płachty na skałach w oceanie. Ich pozostałości można jeszcze dziś zobaczyć w postaci stromatolitów.
Co dzieje się w czasie fotosyntezy i dlaczego jest ona tak ważna?
Cyjanobakterie rozwinęły zdolność wykorzystywania energii światła słonecznego do wiązania CO2 obecnego w środowisku w postaci wysokoenergetycznych związków (cukier!). Te wysokoenergetyczne związki z kolei służą ogromnej większości innych organizmów, w szczególności zwierzętom, a także nam ludziom, jako źródło energii, Spożywamy je jako pożywienie i „spalamy” (przy pomocy tlenu wytwarzanego jako produkt uboczny podczas fotosyntezy) produkując z powrotem CO2. Energia uwolniona w tym procesie napędza wszystkie nasze procesy metaboliczne.
Nawiasem mówiąc, istnieją na to terminy techniczne: cyjanobakterie i wszystkie rośliny wyższe żyją „fotoautotrofowo” (to znaczy czerpią energię ze światła i budują własną biomasę z CO2), wszystkie zwierzęta żyją „chemoheterotrofowo” (zyskują energię z reakcji chemicznych i budują swoją biomasę poprzez przetwarzanie innej biomasy).

Obieg materii i energii na ziemi. W trakcie fotosyntezy z niskoenergetycznych substancji nieorganicznych, przy wykorzystaniu energii słonecznej, powstają bogate w energię substancje organiczne. Te bogate w energię substancje z kolei służą innym organizmom jako pożywienie i dawcy energii.
Dla wszystkich, którzy chcą wiedzieć więcej:
Fotosynteza przebiega w dwóch fazach. W tak zwanej reakcji pierwotnej (zwanej również „reakcją świetlną”) duże kompleksy białkowe w błonach, tak zwane fotosystemy, magazynują energię słoneczną w uniwersalnych cząsteczkach nośnika energii biologicznej: ATP jest znane wielu osobom, ale także tak zwane ekwiwalenty redukcji [H] są nośnikami energii.
- Proste organizmy wykorzystują tylko jeden fotosystem, który jednak nie dostarcza wystarczającej energii do uwolnienia elektronów z wody. Zamiast wody wykorzystuje on na przykład H2S lub proste substancje organiczne jako źródło elektronów. W przypadku tych form fotosyntezy, które zostały już przeprowadzone przez poprzedników cyjanobakterii i które są nadal używane są przez wiele rodzajów bakterii do wytwarzania energii, nie jest uwalniany tlen. Energia tego jednego fotosystemu jest wystarczająca jedynie do wytworzenia równoważników redukcji, które można wykorzystać do dalszych biosyntez.
- W całkowicie wyewoluowanej, tlenowej fotosyntezie cyjanobakterii i dzisiejszych roślin lądowych, dwa fotosystemy są połączone szeregowo — jak silnik z turbodoładowaniem lub pociąg z dwiema lokomotywami. Dopiero drugi fotosystem dostarcza energii wystarczającej do rozszczepienia wody na wodór i tlen. Tlen dyfunduje na zewnątrz, a jony wodoru (H+) są wzbogacane i wykorzystywane do tworzenia ATP.
W reakcji wtórnej (nie jest ona zależna od światła i dlatego jest często mylnie określana jako „reakcja ciemności”), wcześniej wytworzone nośniki energii są wykorzystywane do wiązania cząsteczki CO2 za pomocą specjalnego enzymu na cząsteczce cukru C5, który jednocześnie rozpada się na dwie cząsteczki cukru C3. Z nich można następnie wyprodukować dalsze cząsteczki cukru z cząsteczkami zawierającymi 3, 4, 5, 6 lub 7 atomów węgla, a z tych cukrów wszystkie elementy budulcowe potrzebne organizmowi; kolejne cukry, tłuszcze, białka.
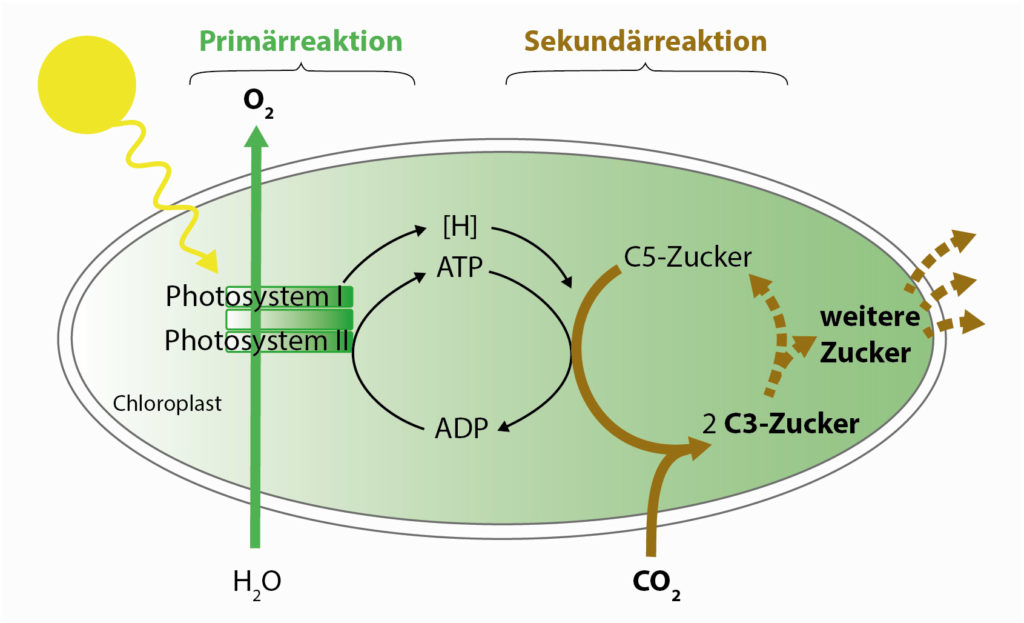
Dwa etapy fotosyntezy: W reakcji pirwotnej energia światła słonecznego jest magazynowana w biochemicznych nośnikach energii (ATP i [H]). W reakcji wtórnej energia ta jest wykorzystywana do wiązania cząsteczki CO2 z cząsteczką 5‑cukrową. Z kolei z powstałego 3‑cukru może powstać wiele innych cząsteczek życia.
Życie bez i z fotosyntezą
Jak wiadomo, życie na Ziemi istniało jeszcze przed rozwojem fotosyntezy. Te bardzo wczesne formy życia wykorzystywały stosunkowo niskie gradienty energii prostych reakcji fizykochemicznych do napędzania swojego metabolizmu (mówiąc językiem technicznym: żyły chemoautotroficznie). Zdolność do fotosyntezy, innymi słowy do wykorzystania energii zawartej w świetle słonecznym, dała jego „wynalazcom” olbrzymią przewagę nad wczesnymi formami życia.
A życie takie, jakie istnieje obecnie na Ziemi, nie byłoby w ogóle możliwe bez fotosyntezy tlenowej.
- Uformowane fotosyntetycznie cząsteczki służą jako pokarm dla wszystkich organizmów wyższych. Używają ich one jako elementów budulcowych, a dzięki zawartej w nich energii napędzają własny metabolizm.
- Bez fotosyntezy brakowałoby nie tylko organicznych cegiełek budulcowych, ale także tlenu do ich wydajnego rozkładu.
My, ludzie, potrzebujemy tlenu do naszego metabolizmu, jesteśmy organizmami tlenowymi. Podobnie wszystkie zwierzęta i rośliny. Wiele bakterii i mikroorganizmów (tzw. beztlenowców) prowadzi metabolizm bez tlenu, zastępując go np. azotem lub siarką. - I wreszcie, rozwój dzisiejszego tak różnorodnego życia nie byłby możliwy bez ochronnej warstwy ozonowej w zewnętrznej atmosferze. Ozon (O3) pochłania dużą część promieniowania UV, które jest szkodliwe dla wszystkich żywych istot, a także oczywiście istnieje dopiero od chwili, gdy wolny tlen znalazł się w atmosferze.
- Nawiasem mówiąc, wszystkie paliwa kopalne, których używamy dzisiaj jako „napęd” naszych niebiologicznie przebiegających procesów lub do wytwarzania ciepła (np. węgiel brunatny, węgiel kamienny, ropa naftowa, gaz) są produktami ubocznymi fotosyntezy.
Jednak fotosynteza, podobnie jak wszystkie formy życia w ogóle, była początkowo ograniczona do oceanów przez ponad 2 miliardy lat i była prowadzona przez cyjanobakterie pływające swobodnie w wodzie lub osadzone na skałach w płytkiej wodzie. Dopiero wraz z rozprzestrzenianiem się wczesnych roślin poza linię wody (początkowo w postaci rozległych mat na kamieniach na obrzeżach pływów, a znacznie później, gdy pierwsze rośliny wyszły na brzeg), pojawiło się życie na lądzie i fotosyntetyczna wymiana gazowa odbywała się bezpośrednio w atmosferze.
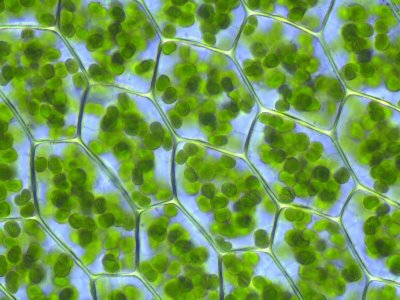
Chloroplasty w liściach mchu liściastego: Chloroplasty to aktywne fotosyntetycznie organelle komórkowe dzisiejszych alg i roślin. Są wynikiem endosymbiozy cyjanobakterii. (Zdjęcie: Kristian Peters, z wikipedii, artykuł „Fotosynteza”. Licencja: GNU FDL 1.2.)
Skąd wiemy, kiedy rozwinęła się fotosynteza?
Na początek: nie wiemy tego dokładnie, ale nadal są intensywne badania. Fotosynteza mogła się rozpocząć już 3,8 miliarda lat temu, prawdopodobnie ponad 3 miliardy lat temu, a prawie na pewno najpóźniej 2,7 miliarda lat temu.
Znaleziska skamieniałości cyjanobakterii
Skamieniałości wspomnianych już stromatolitów, które znajdujemy w warstwach liczących 3,4 miliarda lat, świadczą o ich bardzo wczesnym powstaniu. Z pewnością istniały 2,7 miliarda lat temu,gdyż tyle mają najstarsze skamieniałe cyjanobakterie, jakie do tej pory znaleziono.
Wykrywalne konsekwencje wzrostu stężenia tlenu
Niemal ważniejsze od tych skamieniałości dla klasyfikacji czasowej są zmiany, jakie tlen wytworzył się w trakcie fotosyntezy tlenowej spowodował w jego środowisku i które można wykryć do dziś.
Aż do 3 miliardów lat temu w ziemskiej atmosferze były tylko śladowe ilości tlenu. Powstał, gdy promieniowanie UV rozszczepiło wodę w zewnętrznych warstwach atmosfery. Wodór, który również został wyprodukowany, umknął w kosmos, a (cięższy) tlen został zatrzymany w atmosferze przez grawitację.
Wraz z rozwojem pierwszych organizmów fotosyntetyzujących zawartość tlenu w atmosferze powoli rosła. Można to wykazać za pomocą różnych izotopów chromu, które są utleniane w różnym stopniu. 53Cr znacznie łatwiej się utlenia niż 52Cr. Podczas gdy utleniony chrom jest łatwo wypłukiwany ze skały, nieutleniony chrom pozostaje związany w skale. Analiza warstw skalnych sprzed 3 do 2,4 miliarda lat pokazuje, że w tym czasie udział 53Cr w skałach kontynentalnych spadł, podczas gdy w oceanicznych skałach osadowych wzrósł. Jest to wskazówka na rosnące stężenie O2 w atmosferze, co prowadzi do utleniania 53Cr i jego wymywania.
Zawartość tlenu w oceanach zmieniała się tylko powoli w porównaniu do zawartości w atmosferze. Tlen początkowo utleniał Fe2+, który znajdował się wtedy w dużych ilościach w oceanach. W ten sposób powstało nierozpuszczalne Fe3+ (w postaci Fe2O3 lub tlenku żelaza), które osiadały jako rdza na dnie morskim. Te złoża żelaza w warstwach skalnych sprzed około 2,7 miliarda lat są nadal zachowane jako tak zwane rudy pasmowe, obecnie znajdujące się w większości już nie pod dnem morskim, ale osadzone w warstwach skalnych, z których są wydobywane przez ludzi i stanowią podstawę naszej industrializacji (ponad 90% zużywanego ekonomicznie żelaza pochodzi z pokładów rudy zdeponowanych 2,7 miliarda lat temu!).

Ruda wstążkowa z typowym rozwarstwieniem minerałów tlenku żelaza i kamienistych osadów. Zdjęcie: André Karwath.
CC BY-SA 2.5Uwagi dotyczące dalszego użytkowania; Plik: Black-band ironstone (aka) .jpg; Utworzono: 26 sierpnia 2005 r.
Lokalizacja: 51° 2′ 34,84″ N, 13° 45′ 26,67″ E
.jpg¶ms=51.043011_N_13.757408_E_&language=de){kind=link}
Tlen – zarówno trucizna jak i dawca życia
Przez długi czas stężenie wolnego tlenu w wodzie i atmosferze zmieniało się tylko powoli, ponieważ został on ponownie utrwalony w postaci tlenków metali prawie w takim samym stopniu, w jakim się wytworzył. Jednak gdy metale, zwłaszcza Fe2+, zostały zużyte, stężenie tlenu zaczęło wzrastać niezwykle szybko.
Jest bardzo prawdopodobne, że wczesne, organizmy sciśle beztlenowe nie radziły sobie z tlenem pojawiającym się nagle w ich środowisku, gdyż jest on wysoce toksyczny dla form życia, które nie są przystosowane do O2. W ich metabolizmie prowadzi do powstania wysoce reaktywnych nadtlenków, które uszkadzają ważne elementy strukturalne (błony, białka). Większość beztlenowych form życia, które istniały do tej pory, wymarła. Dopiero rozwój peroksydaz, enzymów, które mogą rozkładać szkodliwe nadtlenki, umożliwił organizmom przetrwanie w obecności tlenu.
Innym czynnikiem, który przyczynił się do „Wielkiej katastrofy tlenowej”, było utlenianie atmosferycznego metanu (CH4) do CO2 i wody. Metan jest znacznie silniejszym gazem cieplarnianym niż CO2. Spadające stężenie metanu było jedną z przyczyn ekstremalnej epoki lodowcowej („Ziemia jak kula śnieżna”), która trwała od 300 do 400 milionów lat i prawdopodobnie spowodowała wymarcie wielu istniejących w tym czasie prymitywnych form życia.
2,2 miliarda lat temu zawartość tlenu w atmosferze ponownie znacznie wzrosła (do prawie jednej piątej dzisiejszej zawartości, czyli do około 3,8%). Przypuszcza się, że ma to związek z końcem fazy oblodzenia: erozja lodowca zmyła zapasy minerałów do oceanów, a CO2 niesiony do atmosfery przez erupcje wulkanów dostarczył materiału niezbędnego do ogromnego wzrostu liczby organizmów aktywnych fotosyntetycznie. Rezultatem był pierwszy na świecie „rozkwit glonów”. W tym czasie rozległe biofilmy pokrywały kamienie w płytkiej wodzie. Przy zawartości tlenu 3% został osiągnięty próg, który doprowadził do powstania warstwy ozonowej w stratosferze. Powstała bariera ochronna przed wrogiegim promieniowaniem UV, która umożliwiła dalszy rozwój i rozprzestrzenianie się życia na lądzie. Około 350 milionów lat temu zawartość tlenu w atmosferze osiągnęła dzisiejszy poziom 21%.
Rozwój metabolizmu tlenowego, czyli układów enzymatycznych, które przy pomocy tlenu rozkładają biocząsteczki, a tym samym uwalniają znacznie więcej energii (oddychanie komórkowe) niż można uzyskać w procesach beztlenowych, ostatecznie umożliwił ewolucję wielu różnorodnych wyższych form życia. Wszystkie wielokomórkowe formy życia na Ziemi uzależnione są od tlenu jako czynnika utleniającego (Jedynym wyjątkiem jest mniej niż jednomilimetrowy organizm z grupy Loricifera, który został odkryty zaledwie kilka lat temu w osadzie dennym Morza Śródziemnego). Praktycznie wszystkie organizmy, które są trwale zdolne do życia bez tlenu, są jednokomórkowe, Większość z nich to bakterie i archeony, tylko kilka z nich to jednokomórkowce eukariotyczne, mające również jądro komórkowe. [MM]