







1300 milionów lat przed dniem dzisiejszym
![]()
Epoka: proterozoik / ektas
 Pod koniec tej epoki po raz pierwszy pojawiają się komórki, które w przeciwieństwie do bakterii mają prawdziwe jądro, w którym przechowywany jest ich materiał genetyczny. Przez to materiał genetyczny jest chroniony. Podział wnętrza komórki na kilka przedziałów umożliwia również oddzielenie różnych reakcji chemicznych i wzbogacenie zapasów substancji.
Pod koniec tej epoki po raz pierwszy pojawiają się komórki, które w przeciwieństwie do bakterii mają prawdziwe jądro, w którym przechowywany jest ich materiał genetyczny. Przez to materiał genetyczny jest chroniony. Podział wnętrza komórki na kilka przedziałów umożliwia również oddzielenie różnych reakcji chemicznych i wzbogacenie zapasów substancji.
Pierwsze komórki tego typu to jednokomórkowe krasnorosty, które później zasiedlają ląd.
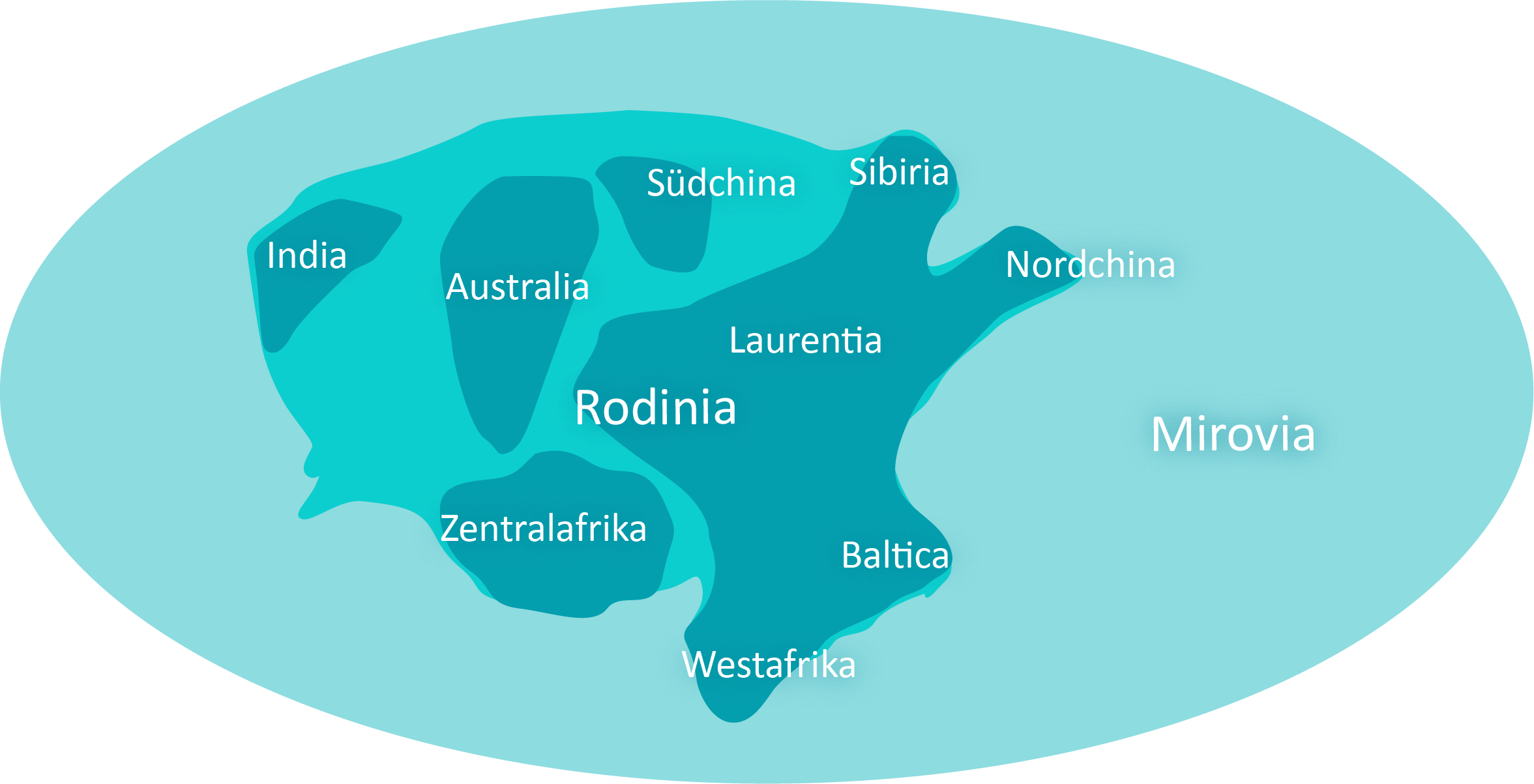
Z geologicznego punktu widzenia rozpoczyna się formowanie superkontynentu Rodinia. Podczas ektasu, który trwa 200 milionów lat, epoki lodowcowe występują wielokrotnie z powodu wyczerpywania się CO2 w atmosferze. Fazy te, w których masy lądowe są gęsto pokryte lodem, przeplatają się z okresami tropikalnymi.
Tę informację w prostym języku znajdziesz na stronie evokids.de.
W ciągu około 2 miliardów lat od powstania pierwszych komórek miały miejsce decydujące wydarzenia dla dalszego życia na Ziemi. W tym niezwykle długim okresie powstały pierwsze komórki z jądrem komórkowym, w którym „upakowany” jest ich materiał genetyczny. Wnętrze komórek podzielone jest przy pomocy bardzo wyraźnych, połączonych ze sobą systemów membranowych oraz zamkniętych organelli na bardzo różne przedziały.
Trzy wielkie królestwa
Obecnie na Ziemi istnieją trzy wielkie królestwa żywych istot: bakterie, archeony i eukarionty (lub eukariota).

3 królestwa żywych istot. Wymieniono tylko najważniejsze grupy. Dwa pierwotne zdarzenia endosymbiozy doprowadziły do rozwoju mitochondriów i pierwotnych plastydów.
Wszystkie organizmy wielokomórkowe należą do eukariota. Oznacza to, że ich komórki mają jądro komórkowe, a także różnią się wieloma innymi właściwościami od dotychczas wyłącznie istniejących archeonów i bakterii, które są zawsze jednokomórkowe i razem tworzą grupę prokariota.
Pierwsza endosymbioza: powstawanie mitochondriów
Istnieją różne hipotezy dotyczące powstawania pierwszych eukariontów, które przez 2 miliardy lat od powstania pierwszych komórek prokariotycznych trwało bardzo długo. Poniżej opisujemy najbardziej prawdopodobną obecnie. Zakłada, że rozwinęła się ścisła symbioza archeonu z bakterią (a dokładniej: α‑proteobakterią). Początkowo nie działo się nic, co nie miało miejsca w wielu wariantach wszędzie tam, gdzie było życie. Komórki (czyli bakterie i archeony) żyły blisko siebie i wymieniały się produktami przemiany materii. Oznacza to, że jeden gatunek wykorzystywał produkty odpadowe innego typu do własnego metabolizmu.Opisany archeon, który później miał się przekształcić się w eukariota, pozyskiwał energię do swoich procesów metabolicznych z reakcji wodoru (H2) z CO2. Metan został uwolniony jako produkt końcowy (jest to znane jako metanogeneza). Potrzebny do tego wodór pozyskiwał początkowo ze źródeł geologicznych, a gdy te wyschły, z wspomnianych powyżej sąsiednich proteobakterii, które również dostarczały CO2. Z kolei proteobakterie przekształcają związki organiczne (ich „substraty” lub pokarm) w CO2 i wodór w trakcie procesów fermentacyjnych.
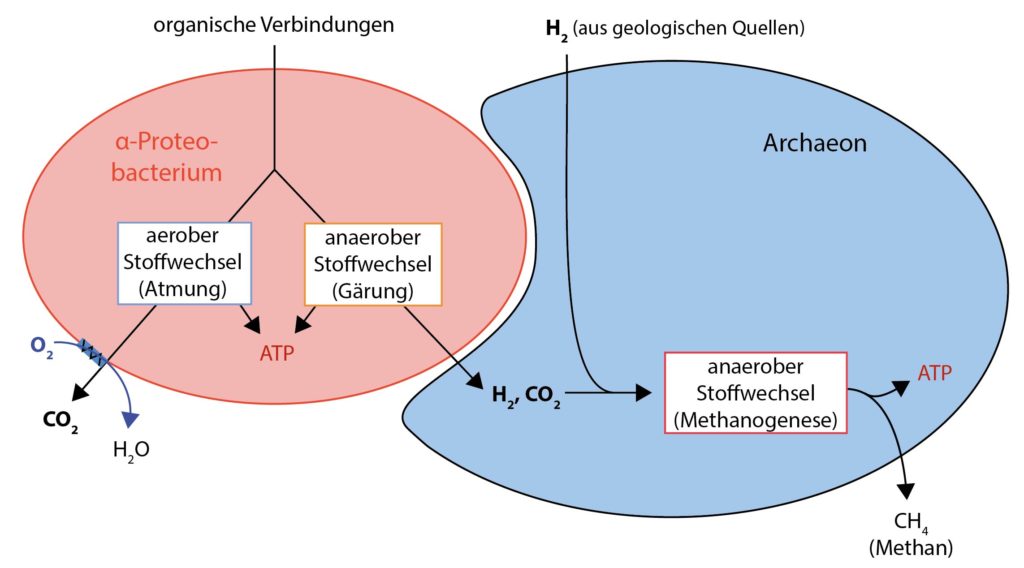
Endosymbioza bakterii przez archeona (hipoteza wodorowa). Archeon czerpie korzyści z metabolizmu bakterii, co uniezależnia go od geologicznych źródeł H2. Ponadto archeon stricte beztlenowy wykorzystuje bliską symbiozę z bakterią, ponieważ powoduje to redukcję tlenu obecnego w środowisku do wody.
Podczas gdy archeon żył w warunkach ściśle beztlenowych, proteobakterie były fakultatywnie beztlenowe, co oznacza, że chociaż wolały żyć w środowisku beztlenowym, mogły również przetrwać w obecności tlenu. W warunkach beztlenowych czerpały energię poprzez fermentację, a jeśli tlen był dostępny, używały go do oddychania tlenowego.
Dla archeona symbioza z proteobakterią, oprócz dostarczania jej składników pokarmowych CO2 i H2, była przydatna także pod innym względem, proteobakteria usuwała szkodliwy dla archeona tlen ze środowiska, usuwając go w trakcie swojego oddychania i redukując do wody.
Zysk energii dzięki oddychaniu tlenowemu
Powszechnie rozumiemy, że oddychanie oznacza napełnianie płuc powietrzem poprzez wdychanie i wydychanie. Fachowo nazywa się to „oddychaniem zewnętrznym”. Tutaj jednak przyjrzymy się temu, co jest znane jako „oddychanie komórkowe”. Oznacza to przeniesienie elektronów generowanych podczas metabolizmu (często symbolizowanych jako [H]) do tlenu z utworzeniem wody (H2O). Pod względem energii odpowiada to tak zwanej „reakcji tlenowodorowej”, która uwalnia duże ilości energii. Jak sama nazwa wskazuje, zazwyczaj jest ta reakcja wybuchowa. Z pomocą kilku kompleksów białkowych osadzonych w błonie komórkowej organizmom tlenowym udało się przeprowadzić tę reakcję małymi etapami w bardzo kontrolowany sposób. To pozwoliło im uzyskać jeszcze więcej energii niż w przypadku reakcji fermentacyjnych i utrwalić ją w postaci ATP, uniwersalnej „waluty energetycznej” żywych organizmów.
W tamtym czasie jednak stężenie tlenu było nadal bardzo niskie, większość istniejących wówczas form życia, w tym opisywany archeon, było beztlenowcami.
Oprócz wielu różnic w wyposażeniu metabolicznym Archaea i Bacteria różnią się również budową ścianki komórkowej: Podczas gdy bakterie zawsze mają twardą, ochronną i stabilizującą warstwę zewnętrzną z tak zwanej „mureiny” wokół błony komórkowej, taka bardzo stabilna warstwa u Archaea już nie występuje. Archaea noszą różne, w zależności od gatunku, zwykle cieńsze osłony wokół błony komórkowej, rozpowszechniona jest warstwa z glikoproteiny.
Opisywany archeon, który stał się eukariotem, również znacznie zredukował tę warstwę, tak że jego zewnętrzna powłoka stała się elastyczna. Był w stanie zagnieździć się blisko sąsiednich proteobakterii, zwiększając w ten sposób powierzchnię wymiany z nimi, otaczając je i ostatecznie wchłaniając całkowicie.
W przeciwieństwie do często spotykanej do dziś fagotrofii, czyli owijania się wokół innego organizmu w celu jego strawienia, archeon pozostawił swój symbiont nienaruszony. W ten sposób zostało zapewnione wzajemne zaopatrzenie w produkty przemiany materii, archeon uniezależnił się od abiotycznych źródeł H2, co ogromnie rozszerzyło dostępną mu przestrzeń życiową. Ponadto powstały warunki do radzenia sobie ze stale rosnącym stężeniem tlenu w tym otoczeniu przez kolejne miliony lat.
W następnym okresie materiał genetyczny został przeniesiony z proteobakterii do komórki gospodarza. Przy tym bakteria utraciła zdolność do niezależnego metabolizmu. W ten sposób stała się tak zwanym organellem w dawnej komórce archeonu, czyli pierwotnym mitochondrium. To przejęcie jednego organizmu przez inny nazywa się endosymbiozą.
Mitochondria: elektrownie komórek eukariotycznych
Współczesne eukarioty zyskują bardzo dużo energii dzięki metabolizmowi tlenowemu, dla którego mitochondria mają zasadnicze znaczenie. Tzw. „łańcuch oddechowy” odbywa się na silnie pofałdowanych błonach mitochondrium, czyli jest opisanym powyżej oddychaniem komórkowym. Tutaj, jako ostatni etap metabolizmu, elektrony uwalniane podczas rozpadu związków organicznych są przenoszone do tlenu. Przy tym powsaje woda i energia w postaci ATP.
Wykształcenie się jądra komórkowego
Zaraz na wstępie: jak dokładnie powstało jądro komórkowe, nie jest szczegółowo znane. Pewne jest, że nastąpiła ożywiona wymiana materiału genetycznego między bakteriami tamtych czasów, jak ma to miejsce również obecnie. Jest to znane jako „poziomy transfer genów”. Natomiast transfer materiału genetycznego do komórek potomnych w trakcie podziału / reprodukcji komórki nazywany jest „pionowym transferem genów”.
DNA bakterii i archeonów leży swobodnie w komórce w postaci mniej lub bardziej splątanego dużego pierścienia („chromosom pierścieniowy”). Ponadto występuje zwykle kilka mniejszych pierścieni, tak zwanych plazmidów. Te plazmidy są przekazywane z komórki bakteryjnej do komórki bakteryjnej. Również DNA, które jest obecne w środowisku w bardzo niespecyficzny sposób, może również zostać wchłonięte. Jest ono często, w całości lub częściowo, wchłaniane przez chromosom pierścieniowy, a następnie przekazywane i odczytywane razem z własnymi genami bakterii.
Przypuszcza się, że przed pochłonięciem proteobakteri lub najpóźniej w trakcie endosymbiozy w archeonie wokół własnego materiału genetycznego utworzyła się błona ochronna. To sprawiło, że włączenie obcego DNA do własnego genomu było łatwiejsze do kontrolowania. Przede wszystkim jednak powstało miejsce, w którym przechowywany jest materiał genetyczny, czyli DNA. Tutaj informacje są również tłumaczone na RNA, które jest następnie przetwarzane i transportowane z jądra do cytoplazmy, gdzie zachodzi synteza białek.

Dwie pierwotne endosymbiozy. Endosymbioza proteobakterii przez archeona prowadzi do powstania pierwszego mitochondrium. W tym kontekście błona tworzy się również wokół materiału genetycznego (→ jądro komórkowe). Grzyby i zwierzęta rozwijają się z pierwotnych eukariontów. W drugiej symbiozie wchłaniane są cyjanobakterie (→ chloroplasty), które tworzą protoplastę komórek roślinnych. Obie endosymbiozy są związane z masowym transferem genów z organelli do genomu gospodarza (fioletowe strzałki). Oryginalne ściany komórkowe endosymbiontów i otaczające systemy błon żywiciela są rozpuszczane i zastępowane błonami zbudowanymi z lipidów estrów kwasów tłuszczowych, które są typowe dla współczesnych eukariotów.
Dalsze organelle komórkowe…
W toku ewolucji w komórce eukariotycznej powstało wiele innych organelli, które nie występują u prokariotów i archeonów. W tych systemach błon zachodzą specjalne procesy metaboliczne. Małe pęcherzyki błonowe (po niemiecku zwane Vesikel) są oddzielane od większych systemów i transportują materiały do innych miejsc wewnątrz komórki lub na zewnątrz.
Znanymi przykładami są aparat Golgiego i retikulum endoplazmatyczne bezpośrednio przylegające i połączone z błoną jądrową. Istnieje również wiele innych przedziałów, takich jak lizosomy i peroksysomy, w których zachodzą specjalne reakcje metaboliczne.


Prokariota i eukariota. Źródła: Prokaryot (powyżej): LadyofHats, german text by NEUROtiker — Derivative work of Image:Average prokaryote cell- en.svg by LadyofHats, Gemeinfrei, Link. Eukariote (poniżej): Von Translation by Muellercrtp — Image:Plant cell structure svg.svg from Mariana Ruiz Villarreal. LadyofHats, Gemeinfrei, Link)
{kind=link}
Pod tym linkiem https://www.thinglink.com/scene/565728773106827265 można znaleźć dwie bardzo udane trójwymiarowe i interaktywne reprezentacje komórek prokariotycznych i eukariotycznych.
Ta duża powierzchnia błony wewnętrznej, zwłaszcza mitochondriów, umożliwia przybierać komórkom eukarotycznym o wiele większe rozmiary niż komórki prokariotyczne.
Wielkość komórki
Wielkość, jaką może osiągnąć komórka, zależy od dostarczanych składników odżywczych zarówno z zewnątrz, jak i wewnątrz komórki. Decydującym czynnikiem jest stosunek jej powierzchni do objętości.
Bakterie i archeony zawsze pozostają małe, mają średnio od 1 do 10 µm. Substancje odżywcze są rozprowadzane w komórce na drodze dyfuzji, czyli biernego „napędzania” plazmy komórkowej.
Z jednej strony, komórki eukariotyczne drastycznie zwiększyły swoją metabolicznie aktywną powierzchnię dzięki organellom, wystarczy pomyśleć o niezliczonych fałdach błony, które służą do wytwarzania energii w jednym mitochondrium! Z drugiej strony, dzięki ich cytoszkieletowi, wzdłuż którego między organellami mogą migrować pęcherzyki o różnej zawartości, mają one skuteczny system transportowy do dystrybucji składników odżywczych, cząsteczek sygnałowych, różnego typu białek i produktów przemiany materii w komórce. Dlatego komórki eukariotyczne mogą stać się znacznie większe niż komórki prokariotyczne: ich średnica waha się od 5 µm w komórkach drożdży do 1 mm w olbrzymich amebach i komórkach jajowych. Niektóre komórki eukariotyczne stają się znacznie większe, na przykład ludzkie komórki nerwowe, których wyrostek może dochodzić do 1 m długości. Poszczególne komórki mogą również łączyć się w tak zwane syncytie i stać się bardzo duże lub wydłużone (np. Nasze komórki mięśniowe). Wielojądrzaste komórki roślinne, np. Valonia untricularis, również mają objętość powyżej 1 ml. Niektóre glony kandelabrowe mogą tworzyć rurki komórkowe o długości ponad 10 cm.
…i nowy endosymbiont
Po powstaniu pierwotnego eukarionta (z mitochondriami, systemem błony wewnątrzkomórkowej i jądrem komórkowym) w wyniku pierwszej opisanej powyżej endosymbiozy, w toku ewolucji wystąpiła druga endosymbioza. W tym przypadku oryginalny eukariot wchłonął cyjanobakterię i stał się to prekursorem dzisiejszych chloroplastów i innych plastydów. Tak powstał pierwszy protoplasta dzisiejszych komórek roślinnych. Większość genów oryginalnego plastydu została również z czasem przeniesiona do jądra komórkowego i zintegrowana z genomem jądrowym. Transfer genów w przebiegu endosymbiozy doprowadził do powstania chromosomów, które do dziś charakteryzują wszystkie genomy eukariotyczne.
Porównania sekwencji DNA i rRNA z dzisiejszych eukariontów pokazują, że te dwa pierwotne zdarzenia endosymbiozy według wszelkiego prawdopodobieństwa wystąpiły tylko raz w toku ewolucji. Wszystkie dzisiejsze mitochondria i wszystkie dzisiejsze plastydy pochodzą więc od jednego wspólnego przodka.
W dalszym przebiegu ewolucji doszło do endosymbiozy komórek, które już zawierały plastydy (endosymbioza wtórna). Przykłady tego można znaleźć w Euglenophyta i Chlorarachniophyta. Występowały również trzeciorzędowe endosymbiozy, w których haptomonady z wtórnymi plastydami były połykane przez bruzdnicowate. W ten sposób powstały warianty plastydów z trzema lub więcej membranami.
Skąd wiemy, kiedy miały miejsce te wydarzenia?
Najstarszymi wiarygodnymi dowodami na obecność eukariotów są dziś mikroskamieniałości datowane na 1,5 miliarda lat temu.
Biogeochemiczne ślady pewnych cząsteczek tłuszczu typowych dla eukariotów w znacznie starszych warstwach, które sugerowałyby wcześniejsze powstanie, okazały się zanieczyszczeniami. [MM]