







560 Millionen Jahre vor heute
![]()
Zeitalter: Neoproterozoikum / Ediacarium
 Aus dieser Zeit stammen die ersten Fossilien von Anhäufungen miteinander verbundener Zellen. Durch die Verbindung legen sie die Grundlage für die Arbeitsteilung unter den Zellen und die Entwicklung von komplexeren Lebewesen.
Aus dieser Zeit stammen die ersten Fossilien von Anhäufungen miteinander verbundener Zellen. Durch die Verbindung legen sie die Grundlage für die Arbeitsteilung unter den Zellen und die Entwicklung von komplexeren Lebewesen.
Strukturproteine geben den sich rasch entwickelnden Lebewesen Form und Halt. Durch Einlagerung von Kalk in die Skelette entstehen feste Panzer, die die Organismen nicht nur stützen, sondern auch Schutz gegen Verletzungen bieten. Damit können größere Lebensformen entstehen. Viele grundlegende Baupläne von Lebewesen entstehen und existieren noch heute.
Schwimmend ist der Trilobit Cheirurus ingricus zu sehen, am Boden ein Exemplar von Dickinsonia.
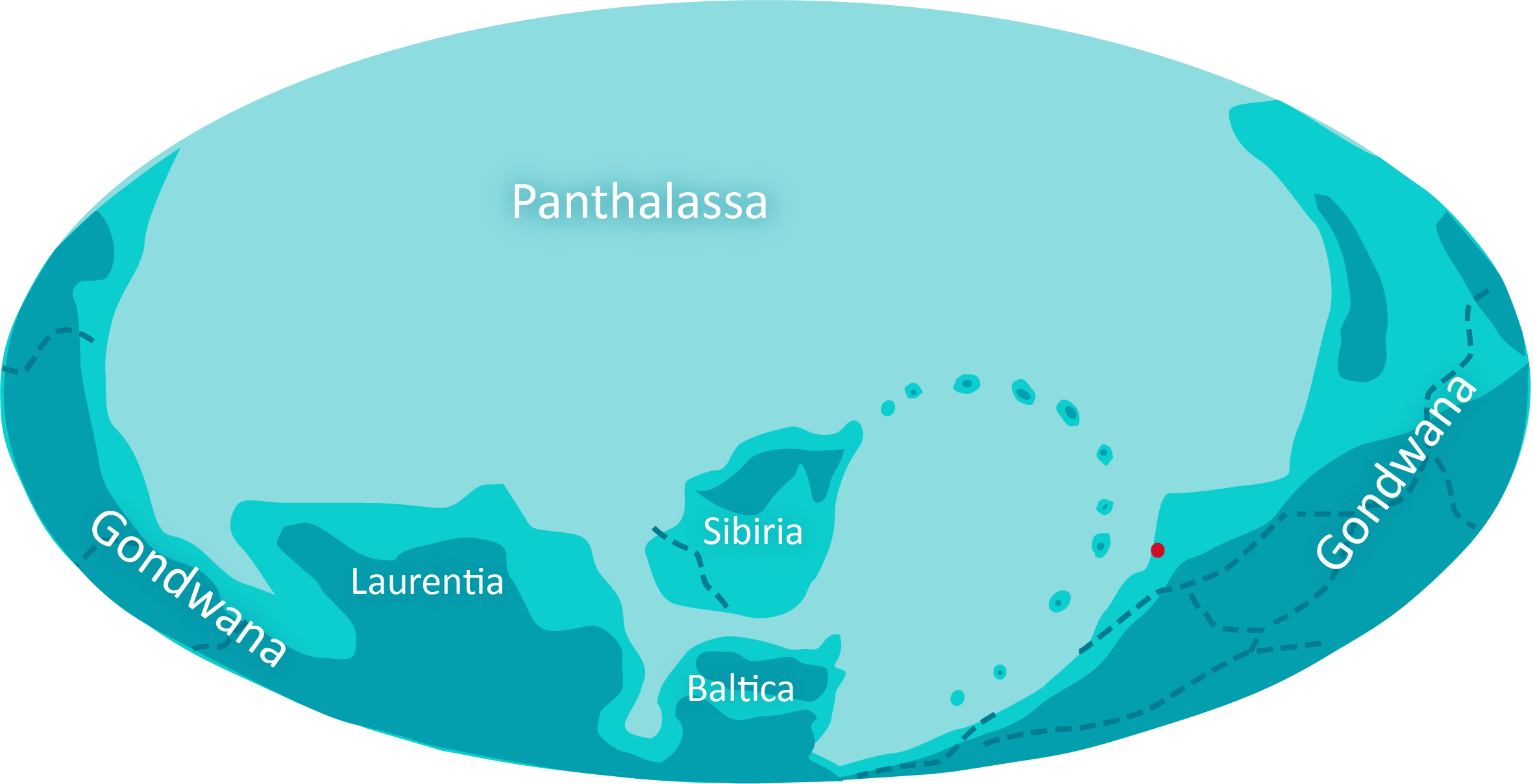
Die Verschiebung von Landmassen in Polnähe unterbricht den Zyklus aus abwechselnder Vereisung und Überhitzung der Erde. Der CO2-Gehalt der Atmosphäre stabilisiert sich und damit auch die Temperatur.
Diese Infos in einfacher Sprache auf evokids.de.
Diese Station im Hörspiel:
Zu den Stütz- und Schutzskeletten zählt man heute Exoskelett, Endoskelett und Hydroskelett.
Das älteste bisher als Fossil gefundene Tier mit Stützskelett ist Namapoikia reitogensis. Ob es sich dabei um eine Koralle oder einen Schwamm handelte ist unsicher, sein Alter wird auf ca. 550 Mio. Jahre bestimmt.
Als die ersten größeren vielzelligen Organismen vor ca. 560 Mio. Jahren entstanden, konnte dies nur gleichzeitig mit der Entwicklung und Entstehung von Stütz- und Schutzskeletten geschehen, die sowohl im Tier- wie auch im Pflanzenreich stattgefunden hatte. Während sich bei den Pflanzen als wichtigstes Stützmolekül das Lignin entwickelte, das neben mineralischen Einlagerungen zwischen die Zellulosefasern der Außenwände eingebaut wurde, hat sich bei vielen Tieren für die äußere Stabilisierung des Körpers das Chitin entwickelt. Chitin ist, wie die Zellulose, ein Polysaccharid, allerdings aus Monomeren von N‑Acetylglucosamin aufgebaut; dieses besteht ebenfalls aus Glucose, trägt jedoch noch einen acetylierten Aminrest.
Chitin stammt wohl ursprünglich von den Bakterien, bei denen es Hauptbestandteil der Zellwand ist.
Warum wachsen Zellen (wenn sie können)?
Zellen werden größer, weil es thermodynamisch, also physikalisch/energetisch in vielen Fällen vorteilhaft ist. Das Verhältnis zwischen Volumen und Oberfläche einer Zelle ist ein Drittel des Kugelradius:

Das heißt, je größer eine Kugel wird, umso stärker wächst ihr Inhalt gegenüber ihrer Oberfläche. Damit sind Wasser‑, Stoff- und Wärmeaustausch mit der Umgebung besser kontrollierbar. Dieses Phänomen der Größenzunahme ist bei vielen Einzellern zu beobachten, die unter limitierter Nährstoffzufuhr leben. Allerdings hat eine Volumenzunahme auch Nachteile, z. B. werden die auftretenden Kräfte bei osmotischen Druckdifferenzen zwischen Zellinnerem und ‑äußerem deutlich größer. Abhilfe schafft hier ein mechanischer Schutz. Zellen ohne einen solchen Schutz können nur im osmotischen Gleichgewicht mit ihrer Umgebung existieren und sind zudem empfindlich gegen mechanische Scherkräfte.
Bereits die Einzeller, prokaryotische wie eukaryotische, haben deshalb schon in einem frühen Stadium der Evolution eine Zellwand aus Stützproteinen und ‑polysacchariden, sogenannten Peptidoglykanen, entwickelt, die ihren Zellen mechanischen Schutz bietet. Dies geschah bereits vor mindestens 3,5 Milliarden Jahren, wie Fossilfunde von Cyanophyceenvorläufern zeigen. Einzige Ausnahme sind parasitisch lebende Bakterien wie Mycoplasmen u.ä.
Für größere Zellverbände und vielzellige Organismen ist eine solche Zellwand allerdings nicht mehr ausreichend. Erst die Bildung von weiteren Stabilisierungsstrukturen löste dieses Problem.
Exoskelett
Die früheste dieser Strukturen war die Zellulose, die aus langen Ketten reiner Glucosemoleküle (Traubenzucker) in spezieller chemischer Bindung besteht. Diese Zellulose wird in der Plasmamembran der Zellen gebildet, nach außen abgeschieden und gleich vernetzt. Mit einer einzigen Ausnahme, den Manteltieren (Tunicata), sind nur Pflanzenzellen zur Synthese und Sezernierung (Ausscheidung) von Zellulose befähigt. Durch ihre Struktur ist die Zellulosefaser sehr gut zugbelastbar, hält aber kaum Drücken stand (Druckstabilität erreichten die Pflanzen erst durch Einlagerung von Lignin zwischen die Zellulosefasern).
Im Tierreich hat sich das Exoskelett außer bei den genannten Ausnahmen speziell bei den Gliederfüßern (Arthropoda) als äußere Abscheidung der Epidermis gebildet. Sie wird Cuticula genannt und besteht als wesentliches Strukturelement aus farblosen, biegsamen Chitinfibrillen. Eine Proteinkomponente, das bräunliche Sklerotin, umhüllt diese Fibrillen und bestimmt die Festigkeit des Komplexes. Als zusätzliche Verfestigung können auch Minerale wie Calcit oder Apatit eingelagert werden, aber auch Zink- oder Magnesiumverbindungen. An den einzelnen Elementen des Außenskeletts setzen von innen Muskeln an, die es ermöglichen, die Elemente gegeneinander zu bewegen.
Da Außenskelette nicht auflösbar sind und auch nicht mitwachsen, sind die Arthropoden gezwungen, diesen Panzer während der Wachstumsphasen zu verlassen und ein neues größeres Außenskelett zu bilden. Dieser Vorgang wird „Häutung“ genannt. Da der Chitinpanzer keinen Schutz gegen Wasserverlust darstellt, ist bei vielen Gliederfüßern auf die Cuticula eine wachsartige Schicht aufgelagert, die Epicuticula genannt wird.
Hydroskelett
Phylogenetisch älter als das Exoskelett, also in der Evolution noch vor diesem entstanden, ist das Hydroskelett. Von einem Hydroskelett spricht man, wenn ein Teil der Muskulatur gegen ein prall gefülltes Flüssigkeitspolster arbeitet, so dass mechanische Kräfte aufgenommen und effektiv weitergeleitet werden können, ähnlich wie bei einem technischen Hydrauliksystem. Dieses System ist vermutlich bei den Faden- und Ringelwürmern entstanden und existiert dort bis heute.
Aber auch Insekten nutzen dieses Hydroskelett trotz ihres ausgeprägten Exoskeletts z. B. für die Funktion ihrer Spring- oder Laufbeine und auch, um sich während der Häutungsvorgänge ihres Exoskeletts zu entledigen. Auch der sogenannte Zellturgor der Pflanzen, bei dem der gegenüber dem Außenmedium erhöhte osmotische Druck des Zellinneren zu einer hydrostatischen Druckdifferenz führt, die von der Zellwand aufgefangen werden muss, entspricht funktionell einem Hydroskelett.
Endoskelett
Das Endoskelett oder Innenskelett ist das auffälligste gemeinsame Merkmal der Wirbeltiere. Es handelt sich dabei um eine mechanische Stützstruktur (Skelett), die nicht Teil der äußeren Hülle des Organismus ist. Seine einzelnen Elemente werden Knochen genannt. Während bei den Arthropoden die Muskeln innen am Außenskelett ansetzen, setzen sie bei den Wirbeltieren an den im Körperinneren gelegenen Knochen an und bewegen die einzelnen Körperteile gegeneinander. Mit Ausnahme der Knorpelfische, deren Endoskelett aus Knorpel mit eingelagertem Calciumcarbonat bestehen, ist das Endoskelett der meisten Wirbeltiere aus Calciumphosphat und Kollagen aufgebaut. Weiterer Bestandteil des Innenskeletts ist Knorpel. Im Gegensatz zu einem Außenskelett aus Chitin wächst ein solches knöchernes Innenskelett mit dem Organismus mit: Jeder Knorpel und auch jeder Knochen unterliegt einem ständigen Abbau und Wiederaufbau. [KHB]