







350 Millionen Jahre vor heute
![]()
Zeitalter: Paläozoikum / Karbon
 Pflanzen bilden die stabile Substanz Lignin als Stützgerüst und zum Schutz. Dieser Holzstoff ermöglicht sehr hohe Wuchsformen. Er kann noch für eine lange Zeit nicht von Mikroorganismen abgebaut werden. Die Pflanzen verfaulen nicht, sondern werden zu Kohle. Der CO2-Gehalt der Luft sinkt, der Sauerstoffgehalt steigt. Deshalb können Insekten sehr groß werden und die neuen Wälder besiedeln.
Pflanzen bilden die stabile Substanz Lignin als Stützgerüst und zum Schutz. Dieser Holzstoff ermöglicht sehr hohe Wuchsformen. Er kann noch für eine lange Zeit nicht von Mikroorganismen abgebaut werden. Die Pflanzen verfaulen nicht, sondern werden zu Kohle. Der CO2-Gehalt der Luft sinkt, der Sauerstoffgehalt steigt. Deshalb können Insekten sehr groß werden und die neuen Wälder besiedeln.

Euramerika (Laurussia) und Gondwana kollidieren zu dem neuen Superkontinent Pangäa. In Euramerika herrschen tropische Bedingungen, während die am Südpol liegenden Landmassen von einer zunehmend dickeren Eisschicht bedeckt sind. Durch die Eisbildung sinkt der Meeresspiegel.
Diese Infos in einfacher Sprache auf evokids.de.
Diese Station im Hörspiel:
Die ersten pflanzlichen Lebensformen, die das Land besiedelten, waren schlauchförmig organisiert und erreichten nur niedrige Wuchshöhen. Erst die Entwicklung der Stützsubstanz Lignin (auch Holzstoff genannt), das als druckstabiles Füllmaterial zwischen die zugstabilen Cellulosefasern eingelagert wurde, ermöglichte ihnen, höhere Wuchsformen und schließlich auch selbsttragende Verzweigungen auszubilden. Die Fähigkeit zur Bildung von Lignin, die vor etwa 450 bis 400 Millionen Jahren entstand, war damit ein Schlüsselereignis bei der Eroberung der terrestrischen Lebensräume und prägte fortan ihr Erscheinungsbild.
Das Leben an Land birgt viele Herausforderungen
Die Lebensbedingungen im Wasser waren vergleichsweise einfach: Es filterte einen Teil der schädlichen UV-Strahlung aus dem Sonnenlicht, der Auftrieb ermöglichte fadenförmige oder flächige Wuchsformen, und die Gefahr der Austrocknung war nicht gegeben. Nachdem die ersten Pflanzen an Land gelangt waren, mussten sie sich gegen die Schwerkraft behaupten, durch UV-Licht gebildete, aggressive Substanzen unschädlich machen, der Pflanzenkörper musste vor Wasserverlust geschützt werden, und sie mussten sich neuer Schädlinge und Fraßfeinde erwehren.
Es herrschte also ein gehöriger Evolutionsdruck, der im Laufe der Jahrmillionen zur Ausbildung einer extremen Vielfalt an Stoffwechselleistungen geführt hat. Diese, den allgemeinen Energie- und Baustoffwechsel ergänzenden Prozesse bezeichnet man auch als Sekundärstoffwechsel. Als Sekundärmetabolite wurden beispielsweise Fraßabwehrstoffe und Materialien zum UV- und Verdunstungsschutz gebildet, irgendwann auch Duftstoffe und Farbstoffe zur Anlockung von Bestäubern (s. Blütenpflanzen).
Zu den wichtigsten Errungenschaften zählt dabei der Stoffwechsel des Lignins und seiner Grundsubstanzen.
Lignin bietet vielfältige Vorteile
Schutz vor UV-Schäden: Schon die Einzelsubstanzen, aus denen das Lignin zusammengesetzt ist (Cumaryl‑, Coniferyl- und Sinapylalkohole), sind sehr nützlich: Es handelt sich um sogenannte Phenylpropanoide, das sind mit konjugierten Doppelbindungen versehene Kohlenwasserstoffverbindungen. Substanzen mit solchen Verbindungen bezeichnet man in der Fachsprache als „aromatisch“. Sie „entschärfen“ sogenannte Radikale, also aggressive Moleküle, die durch UV-Einwirkung in allen Zellen entstehen. Auf diese Weise boten sie den ersten Landpflanzen einen Überlebensvorteil, so dass die für ihre Bildung notwendige Enzymausstattung auf zahlreiche Nachkommen vererbt, also evolutionär selektioniert und stabilisiert wurde. Die Enzyme für die Bildung der genannten Bausteine haben sich vermutlich schon vor etwa 450 Millionen Jahren, also in den damals noch im Wasser lebendenden Vorformen der Landpflanzen, entwickelt.
Im Lignin selbst, einem Biopolymer, sind die Einzelbausteine in unterschiedlichen Kombinationen dreidimensional zusammengesetzt. Dadurch entsteht eine sehr stabile Substanz, die hohen Drücken standhalten kann. Durch die vielfältigen chemischen Verbindungstypen zwischen den einzelnen Grundbausteinen ist Lignin auch sehr schwer abbaubar, so dass Fraßfeinden damit viel Widerstand entgegen gesetzt wurde. Die Fähigkeit zur Polymerisierung der Ligninbausteine entstand vor etwa 420 Millionen Jahren. Sie ist kennzeichnend für die Gefäßpflanzen.

Beispiel einer Ligninstruktur (Quelle: Karol Głąbpl.wiki: Karol007commons: Karol007e-mail: kamikaze007 (at) tlen.pl [GFDL (http://www.gnu.org/copyleft/fdl.html), CC-BY-SA‑3.0 (http://creativecommons.org/licenses/by-sa/3.0/) or CC BY-SA 2.5 (https://creativecommons.org/licenses/by-sa/2.5)], via Wikimedia Commons).
Eingelagert in die Zellwand, bildet Lignin in Kombination mit der faserförmigen und biegsamen, aber reißfesten Cellulose eine sehr druck- und zugstabile Struktur – ähnlich wie Stahlbeton, wobei die Rolle der Cellulose mit dem Stahl und die des Lignins mit dem Beton verglichen werden kann.
Eroberung des Landes durch Höhenwachstum
Die Evolution der landlebenden Pflanzen und vor allem der Bäume ist eng mit der Ligninbiosynthese verknüpft: Das Lignin ist erst mit dem Auftreten der sogenannten Gefäßpflanzen (lateinisch: Tracheophyten) als echtes Lignin anzutreffen. Zu den Gefäßpflanzen zählen die Bärlapp-Pflanzen, die Farne und alle Samenpflanzen. In ursprünglicheren Pflanzen, also den meisten Algen- und Moosarten, sind dagegen nur die Bausteine oder Lignin-ähnliche Polymere vorhanden.
Lignin bildet die Kittsubstanz, die den pflanzlichen Zellverbund auch unter Belastung zusammenhält. Es bietet zudem Schutz gegen Eindringen von Wasser in das Zellwandmaterial. Dadurch wurde es den Pflanzen möglich, spezielle Leitgefäße für Wasser und darin gelöste Stoffe zu entwickeln. Das Lignin hält das Wasser in diesen Gefäßen – und natürlich auch in den einzelnen Pflanzenzellen selbst.
Leitgewebe in Pflanzen
Ähnlich wie Tiere Arterien und Venen zum Transport ihrer Körperflüssigkeiten besitzen, sind in den höheren Pflanzen Leitgefäße ausgebildet. Sie sind in der Regel in sogenannten Leitbündeln organisiert. Das Xylem (im oberen Bild die großvolumigen, dunkelrot umrandeten Röhren) dient dem Transport von Wasser und anorganischen Salzen von den Wurzeln durch und in die Pflanze. Im Phloem (im oberen Bild blau) werden Zucker und Aminosäuren von den Orten ihrer Produktion (vorwiegend die Laubblätter) zu den Orten des Verbrauchs (Speicherorgane, wachsende Organe) transportiert.
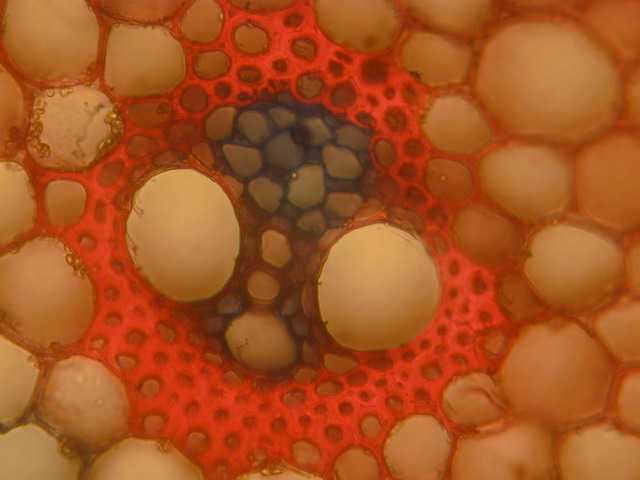
Leitbündel. Als Beispiel ist ein (primäres) Leitbündel der Maispflanze gezeigt. Das Xylem (dunkelrot mit mehreren großlumigen Tracheen) dient dem Transport von Wasser und anorganischen Salzen, im Phloem (blau) werden Zucker und Aminosäuren transportiert. Stabilisiert wird die Struktur durch dickwandiges Stützgewebe (Sklerenchym, hellrot). (Quelle: Von Bamading – selbst fotografiert von Bamading, Gemeinfrei, https://commons.wikimedia.org/w/index.php?curid=798645)

Sekundäres Leitgewebe am Beispiel der Eberesche (Querschnitt durch einen zweijährigen Spross). Das Leitgewebe ist hier konzentrisch organisiert: Das Xylem (rosa) liegt innen, es bildet das Holz. Das Phloem (blau) liegt außen und bildet den Bast. Dazwischen liegt das teilungsaktive Gewebe (Kambium), das das sekundäre Dickenwachstum der verholzenden Pflanzen bewirkt. Die je nach Jahreszeit unterschiedlich großvolumigen Gefäße sind im Holzteil als Jahresringe zu erkennen. PE = Periderm, RP = Rindenparenchym, SK = Sklerenchym – Inseln, PH = Phloem, PMS = primärer Markstrahl, K = Kambium, J = Jahresringgrenze, XY = Xylem. (Urheber: Hans-Jürgen Koch, Weyhe, https://www.mikroskopie-forum.de/index.php?topic=20477.0, mit freundlicher Genehmigung.)
Auch die Ausbildung von Festigungselementen, die bei fehlendem Auftrieb durch das Wasser die Stabilität größerer Pflanzenkörper gewährleisten, und entsprechender, der Schwerkraft entgegen wirkender Tragestrukturen, wurde erst mithilfe des Biopolymers Lignin möglich. Gleiches gilt für die Ausbildung von Verzweigungen und Astsystemen zur Schaffung großer photosynthetisch wirksamer Flächen.
Mit der Fähigkeit zur Bildung von Stützkonstruktionen begann der Siegeszug der Landpflanzen, die in den folgenden Jahrmillionen in vielfältigsten Wuchsformen den terrestrischen Lebensraum besiedelten. Im Wettbewerb um Licht und Wasser entstanden riesige Wälder, zunächst dominiert von Bärlappen und Baumfarnen und schließlich von den heute noch vorherrschenden Samenpflanzen, zu denen auch die Nadel- und Laubbäume (s. Blütenpflanzen) gehören.
Wassertransport gegen die Schwerkraft
Über sogenannte Spaltöffnungen auf der Unterseite der Blätter steht das ansonsten von schlecht wasserdurchlässigen Schichten umhüllte Gewebe der Pflanzen im Austausch mit der Umgebungsluft. Die Spaltöffnungen sind der Ort des Gasaustauschs. Hier wird CO2-reiche Luft aus der Umgebung aufgenommen, um sie der Photosynthese zuzuführen und Baustoffe und sonstige nützliche Substanzen daraus zu bilden. Im Gegenzug wird Sauerstoff, das Abfallprodukt der Photosynthese, an die Umgebung abgegeben. Im Vergleich dazu: Bei uns Menschen erfolgt der Gasaustausch über die Lungenbläschen und mit umgekehrten Vorzeichen: Wir nehmen Sauerstoff auf und geben als Abfallprodukt unseres Stoffwechsels CO2 an die Umgebung ab.
Bei diesem Austausch verliert die Pflanze auch immer Wasser. Das Wasser, das sie an den Blättern verliert, zieht die Pflanze aus ihrem Gefäßsystem nach (genauer: aus dem Xylem). Auf diese Weise entsteht ein Sog, Transpirationssog genannt. Der dadurch ausgelöste Transpirationsstrom dient, neben der Kühlung durch die Verdunstung an den Blättern, vor allem dem Transport von Ionen und Stoffwechselmetaboliten innerhalb der Pflanze.

Um eine Wassersäule zehn Meter entgegen der Schwerkraft anzuheben und zugleich die dabei entstehenden Reibungskräfte des Wassers im Xylem zu überwinden, ist ein Unterdruck von etwa ‑0,3 MPa notwendig. Erst die Einlagerung von Lignin in die Wände des Röhrensystems macht sie stabil genug, dass die Hohlräume dabei nicht kollabieren. Je höher der Baum, desto höhere Unterdrücke werden benötigt, bei Mammutbäumen werden über ‑3 MPa erreicht. Dabei besteht die Gefahr, dass die Wassersäule reißt, weil die Kohäsionskräfte zwischen den Wassermolekülen nicht mehr ausreichen. Dies ist der Grund, warum Bäume nicht beliebig hoch wachsen können, sondern nur maximal etwa 116 m erreichen können. Reines Wasser in Glasröhrchen reißt zwar erst bei etwa 10fach höheren Unterdrücken, allerdings enthält das in Pflanzen transportierte Wasser Ionen und gelöste Gase. Es kommt dadurch sehr viel schneller zu Embolien, also Luftblasen im Wasserstrom, die für die Pflanze – ähnlich wie bei tierischen Organismen – sehr gefährlich sind.
Unterstützt wird der Wasserstrom auch durch osmotische (also auf Verschiebungen von gelösten Substanzen und Ionen beruhende) Kräfte. Auch ohne Transpiration strömt daher Wasser durch die Pflanze, so dass die Transportprozesse gewährleistet sind.
Küstenmammutbaum. Diese Bäume sind mit Wuchshöhen von bis zu 116 m die höchsten Pflanzen auf der Erde. (Quelle: https://de.wikipedia.org/wiki/Riesenmammutbaum#/media/File:General_Grant_tree.jpg)
Lignin: Grundstoff der Kohle
Lignin ist nach der Cellulose und vor dem Chitin die zweithäufigste organische Substanz auf der Erde. Die massive Photosyntheseleistung der das Land erobernden Pflanzen führte im Paläozoikum zu einer immensen Bindung von CO2.
Starben die Pflanzen, fielen sie zu Boden, wo sie Schicht um Schicht von weiteren absterbenden Pflanzen überlagert wurden. Durch die Stabilität der Bindungen im Lignin ist dieses generell sehr schwer abbaubar. Lange konnte es biologisch überhaupt nicht abgebaut werden – die Fähigkeit zum Ligninabbau wurde erst viel später entwickelt. Sauerstoffarme Bedingungen (Sümpfe, Moore) erschwerten den Abbau zusätzlich. Während die meisten anderen Bausteine der toten Pflanzen nach und nach von Mikroorganismen abgebaut wurden, blieb das Lignin mit dem darin enthaltenen Kohlenstoff erhalten. Überlagert von Sedimenten und unter hohem Druck fossilierte es über hunderte Millionen Jahre und wurde zu Steinkohle, die wir noch heute zur Energiegewinnung nutzen.

Steinkohleflöz. Geringmächtige Kohleflöze zwischen „Ruhrsandstein“-Schichten (Sprockhöveler Schichten, Namur C, Oberkarbon) in einem Steinbruch bei Wetter (Ruhr). (By Arnoldius [CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)], from Wikimedia Commons)
Bei der Papierherstellung ist Lignin übrigens unerwünscht, weil es zur Vergilbung beiträgt. Es wird chemisch entfernt und z.B. als Brennstoff oder auch als Grundsubstanz für den Aromastoff Vanillin verwendet. In Lebensmitteln zählt Lignin zu den Ballaststoffen. Es wird vermutet, dass es Karzinogene und andere schädliche Substanzen binden kann und damit unschädlich macht.
Sauerstoffreiche Atmosphäre
Die massive Fixierung von CO2 in Biomasse, die nicht wieder zu CO2 abgebaut wurde, führte zu einem starken Absinken des CO2-Gehalts der Atmosphäre. Parallel stieg der Sauerstoffgehalt an. Am Ende des Karbon lag er bei über 30% (Vergleich heute: 21%). Diese Veränderungen wiederum förderten zahlreiche Innovationen bei anderen Organismen, darunter die Evolution der Flugfähigkeit bei Insekten.
Im Gegensatz zu Wirbeltieren besitzen Insekten keine aktiv be- und entlüfteten Lungen zum Gasaustausch, sondern die Luft gelangt über eher starre Röhrensysteme in den Körper. Da keine aktive Ventilation erfolgt, sondern die Luft mehr oder weniger passiv ein- und ausströmt, ist die Effizienz des Stoffwechsels dieser Tiere in hohem Maße vom Sauerstoffgehalt der Luft abhängig.
Dank der sauerstoffreichen Atmosphäre und des zum Teil tropischen Klimas konnten Insekten und andere Kerbtiere (z.B. Spinnentiere) im Karbon extrem groß werden, wie durch zahlreiche Fossilfunde (z.B. mehr als 2 m lange Tausendfüßler, Libellen mit bis zu 70 cm Spannweite) belegt ist. [MM]

Fossil eines Libellenflügels aus dem Karbon. Dieser etwa 300 Millionen Jahre alte fossile Flügel misst etwa 19 Zentimeter. Zum Vergleich oben links der Flügel der größten Libellenart der letzten 65 Millionen Jahre (Foto von Wolfgang Zessin, auf http://www.sciencemag.org/news/2012/06/where-have-hawk-sized-insects-gone).