







1300 Millionen Jahre vor heute
![]()
Zeitalter: Proterozoikum / Ectasium
 Gegen Ende dieses Zeitalters treten erstmals Zellen auf, die im Gegensatz zu Bakterien einen echten Zellkern besitzen, in dem ihre Erbsubstanz aufbewahrt ist. Dies schützt das Erbmaterial. Die Aufteilung des Zellinnenraums in mehrere Abteile ermöglicht zudem die Trennung verschiedener chemischer Reaktionen sowie die Anreicherung von Speicherstoffen.
Gegen Ende dieses Zeitalters treten erstmals Zellen auf, die im Gegensatz zu Bakterien einen echten Zellkern besitzen, in dem ihre Erbsubstanz aufbewahrt ist. Dies schützt das Erbmaterial. Die Aufteilung des Zellinnenraums in mehrere Abteile ermöglicht zudem die Trennung verschiedener chemischer Reaktionen sowie die Anreicherung von Speicherstoffen.
Zu den ersten Zellen dieser Art zählen einzellige Rotalgen, die später auch das Land besiedeln werden.
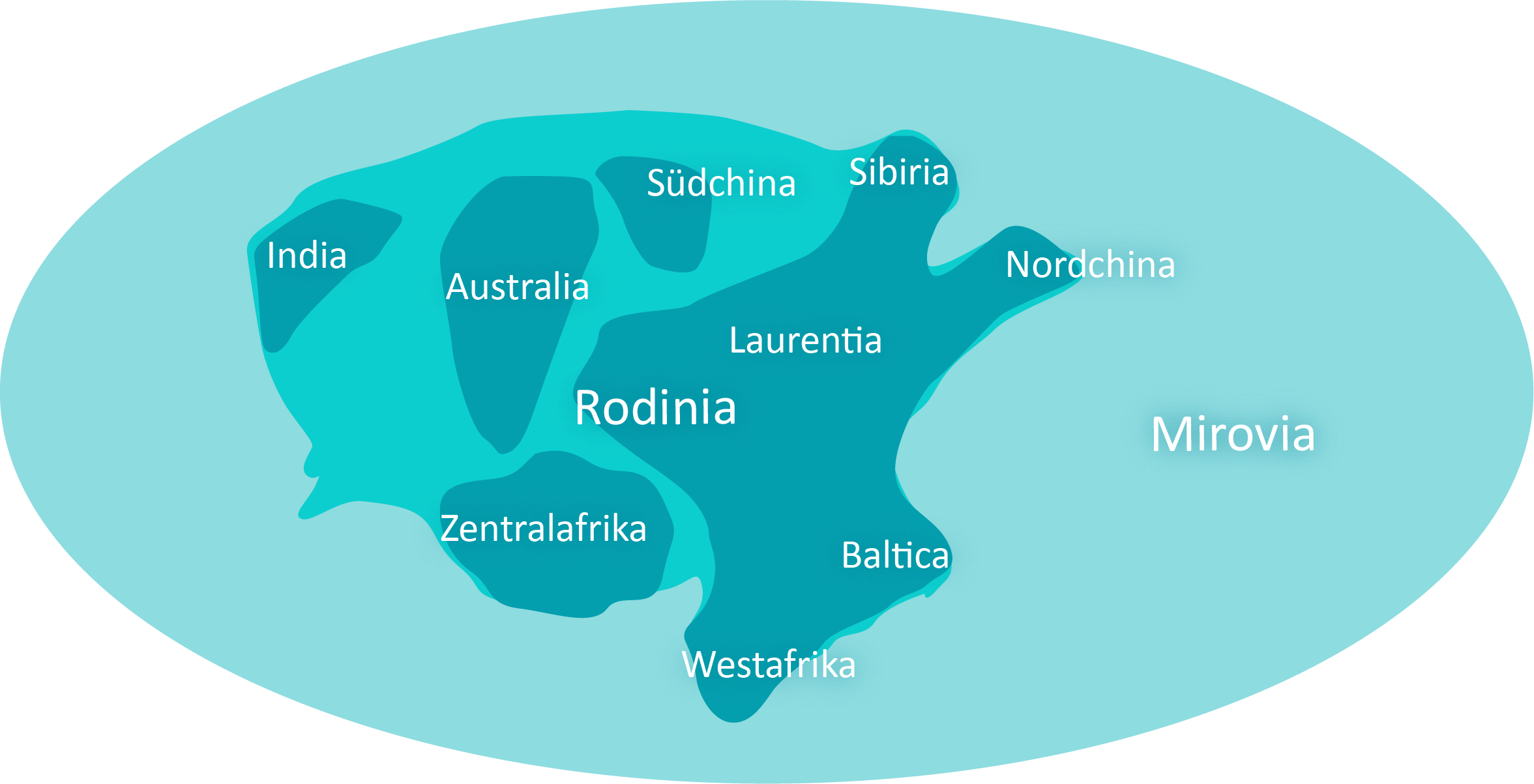
Geologisch beginnt die Bildung des Superkontinents Rodinia. Während des 200 Mio. Jahre dauernden Ectasium kommt es durch CO2-Verarmung der Atmosphäre wiederholt zu Eiszeiten. Diese Phasen, in denen die Landmassen dick mit Eis bedeckt sind, wechseln sich ab mit tropischen Perioden.
Diese Infos in einfacher Sprache auf evokids.de.
Diese Station im Hörspiel:
In den etwa 2 Milliarden Jahren seit der Entstehung der ersten Zellen vollzogen sich für das weitere Leben auf der Erde entscheidende Ereignisse: In dieser extrem langen Zeitspanne entstanden die ersten Zellen mit einem Zellkern, in dem ihre Erbsubstanz „verpackt“ ist und deren Zellinneres durch sehr ausgeprägte, miteinander verbundene Membransysteme sowie abgeschlossene Organellen in sehr unterschiedliche Kompartimente unterteilt ist.
Die drei großen Reiche
Auf der Erde werden heute drei große Reiche von Lebewesen unterschieden: Bacteria, Archaea und Eukarya (oder Eukaryota).

Die 3 Reiche der Lebewesen. Nur die wichtigsten Gruppen sind genannt. Die beiden primären Endosymbiose-Ereignisse führten zur Entstehung der Mitochondrien und der primären Plastiden.
Alle mehrzelligen Organismen gehören den Eukaryota an. Das heißt, ihre Zellen besitzen einen Zellkern und unterscheiden sich auch noch in zahlreichen anderen Eigenschaften von den bis dahin ausschließlich vorhandenen Archaea und Bacteria. Diese sind immer einzellig und bilden zusammen die Gruppe der Prokaryota.
Erste Endosymbiose: Entstehung von Mitochondrien
Zur Entstehung der ersten Eukaryoten, die mit 2 Milliarden Jahren seit der Bildung der ersten prokaryotischen Zellen sehr lang gedauert hat, gibt es unterschiedliche Hypothesen. Wir beschreiben im Folgenden die derzeit wahrscheinlichste. Sie geht davon aus, dass sich zunächst eine enge Symbiose zwischen einem Archaeon und einem Bakterium (genauer: einem α‑Proteobakterium) entwickelt hat. Zunächst geschah dabei nichts, das sich nicht in zahlreichen Varianten allerorts, wo es Leben gab, abspielte: Zellen (also Bacteria und Archaea) lebten in enger Nachbarschaft zueinander und tauschten Stoffwechselprodukte aus. Das heißt, eine Art nutzte die Abfallprodukte anderer Art(en) für ihren Stoffwechsel. „Unser“ Archaeon, das sich später zu einem Eukaryoten entwickeln sollte, gewann die Energie für seine Stoffwechselprozesse aus der Reaktion von Wasserstoff (H2) mit CO2. Als Endprodukt wurde dabei Methan freigesetzt (man bezeichnet dies als Methanogenese). Den Wasserstoff, den es dafür benötigte, erhielt es zunächst aus geologischen Quellen, und als diese versiegten, von den oben erwähnten benachbarten Proteobakterien. Diese lieferten auch das CO2. Denn die Proteobakterien setzten ihrerseits organische Verbindungen (ihre „Substrate“ oder Nahrung) im Rahmen von Gärungsvorgängen zu CO2 und Wasserstoff um.
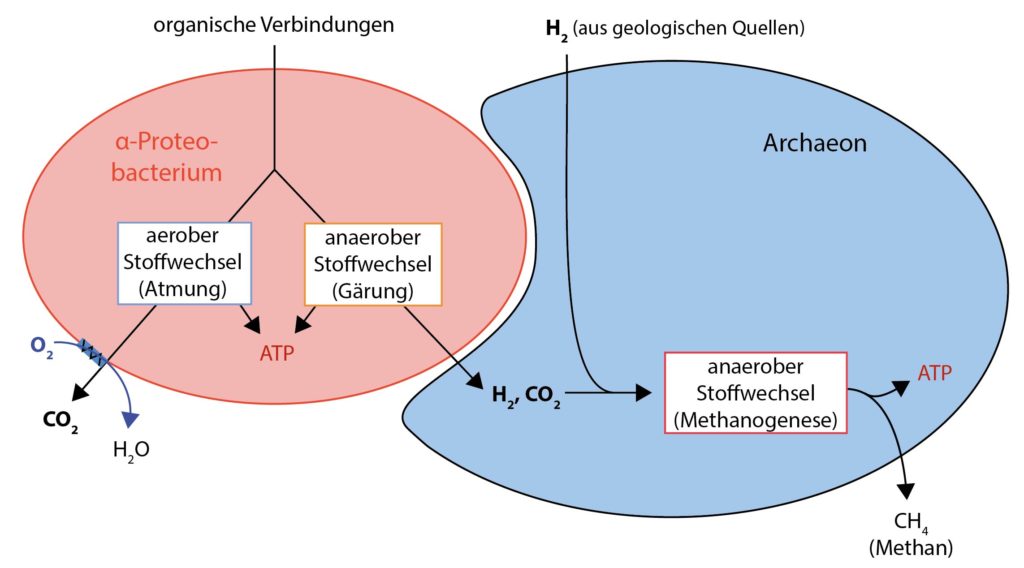
Endosymbiose eines Bakteriums durch ein Archaeon (Wasserstoff-Hypothese). Das Archaeon profitiert vom Stoffwechsel des Bakteriums, durch den es von geologischen H2-Quellen unabhängig wird. Außerdem nutzt dem strikt anaeroben Archaeon die enge Symbiose mit dem Bakterium, weil dieses ggf. in der Umgebung vorhandenen Sauerstoff zu Wasser reduziert.
Während das Archaeon strikt anaerob lebte, war das Proteobakterium fakultativ anaerob, das heißt, es lebte zwar bevorzugt in sauerstofffreier Umgebung, konnte aber auch in Anwesenheit von Sauerstoff überleben. Unter anaeroben Bedingungen gewann es seine Energie durch Gärungen; wenn Sauerstoff vorhanden war, nutze es ihn zur aeroben Atmung.
Für das Archaeon war die Symbiose mit dem Proteobakterium, abgesehen von der Versorgung mit seinen Nährstoffen CO2 und H2, somit auch in einer weiteren Hinsicht nützlich: Das Proteobakterium schaffte den für das Archaeon schädlichen Sauerstoff aus der Umgebung weg, indem es ihn im Zuge seiner Atmung zu Wasser reduzierte.
Energiegewinn durch aerobe Atmung
Landläufig verstehen wir unter Atmung das Befüllen unserer Lungen mit Luft durch Ein- und Ausatmen. Dies nennt man in der Fachsprache „äußere Atmung“. Hier betrachten wir jedoch die sogenannte „Zellatmung“. Darunter versteht man den Transfer der beim Stoffwechsel anfallenden Elektronen (gerne symbolisiert als [H]) auf Sauerstoff unter Bildung von Wasser (H2O). In energetischer Hinsicht entspricht dies der sogenannten „Knallgasreaktion“, bei der sehr viel Energie frei wird. Wie der Name sagt, verläuft sie normalerweise explosiv. Den aeroben Organismen gelang es mithilfe mehrerer, in die Zellmembran eingelagerter Proteinkomplexe, diese Reaktion sehr kontrolliert in kleinen Schritten ablaufen zu lassen. Damit konnten sie noch mehr Energie als mit den Gärungsreaktionen gewinnen und in Form von ATP, der universellen „Energiewährung“ lebender Organismen, fixieren.
Die Sauerstoffkonzentration war zur damaligen Zeit jedoch noch immer sehr gering, die meisten damals vorhandenen Lebensformen, unter anderem „unser“ Archaeon, waren Anaerobier.
Neben vielen Unterschieden in ihren Stoffwechsel-Ausstattungen unterscheiden sich Archaea und Bacteria auch im Aufbau ihrer Zellwand: Während Bacteria immer eine feste, sie schützende und stabilisierende Außenschicht aus sogenanntem „Murein“ um ihre Zellmembran herum besitzen, ist eine solche, sehr stabile Schicht bei den Archaea nicht mehr vorhanden. Archaea tragen je nach Art unterschiedliche, generell dünnere Schutzhüllen um ihre Zellmembran, weit verbreitet ist eine Schicht aus Glykoproteinen.
Unser zum Eukaryoten gewordenes Archaeon hat auch diese Schicht stark reduziert, so dass seine Außenhülle flexibel geworden ist. Es konnte sich eng an ein benachbartes Proteobakterium anschmiegen und seine Austauschfläche mit ihm dadurch immer weiter vergrößern, es umschließen und schließlich komplett in sich aufnehmen.
Im Gegensatz zur auch heute noch häufig anzutreffenden Phagotrophie, also dem Umschlingen eines anderen Organismus, um ihn anschließend zu verdauen, ließ das Archaeon seinen Symbionten allerdings intakt. Auf diese Weise war die gegenseitige Versorgung mit Stoffwechselprodukten gesichert, das Archaeon wurde von abiotischen H2-Quellen unabhängig, was seinen erschließbaren Lebensraum enorm erweitert hat. Außerdem waren die Voraussetzungen geschaffen, um mit der in den folgenden Jahrmillionen stets ansteigenden Sauerstoffkonzentration in der Umgebung zurecht zu kommen.
In der Folgezeit kam es zu einem Transfer von Genmaterial von dem Proteobakterium in die Wirtszelle. In diesem Zuge verlor das Bakterium die Fähigkeit zu einem eigenständigen Stoffwechsel. Es ist damit zu einem sogenannten Organell innerhalb der ehemaligen Archaeon-Zelle geworden, nämlich zu einem Ur-Mitochondrium. Diese Aufnahme eines Organismus durch einen anderen bezeichnet man als Endosymbiose.
Mitochondrien: Die Kraftwerke eukaryotischer Zellen
Moderne Eukaryoten gewinnen sehr viel Energie mit ihrem aeroben Stoffwechsel, für den die Mitochondrien von essenzieller Bedeutung sind: An den stark gefalteten Membranen des Mitochondriums findet die sogenannte „Atmungskette“ statt, d.h. die oben bereits beschriebene Zellatmung. Hier werden, als Endstrecke des Stoffwechsels, die beim Abbau von organischen Verbindungen freiwerdenden Elektronen auf Sauerstoff übertragen. Dabei entstehen Wasser und Energie in Form von ATP.
Herausbildung des Zellkerns
Gleich vorweg: Wie der Zellkern genau entstanden ist, weiß man nicht im Detail. Sicher ist, dass es zwischen den Bakterien der damaligen Zeit, ebenso wie bei heutigen Bakterien, einen regen Austausch von Genmaterial gab. Man bezeichnet dies als „horizontalen Gentransfer“. Dagegen bezeichnet man die Übertragung von Erbmaterial auf die Tochterzellen im Zuge der Zellteilung/Vermehrung als „vertikalen Gentransfer“.
Die DNA von Bakterien und Archaeen liegt frei in der Zelle, und zwar in Form eines mehr oder weniger verknäuelten großen Ringes („Ringchromosom“). Daneben gibt es meist noch mehrere kleinere Ringe, sogenannte Plasmide. Diese Plasmide werden von Bakterienzelle zu Bakterienzelle weitergegeben. Es kann aber auch ganz unspezifisch in der Umgebung vorhandene DNA aufgenommen werden. Diese wird häufig, ganz oder teilweise, ins Ringchromosom integriert und dann gemeinsam mit den bakterieneigenen Genen weitergegeben und abgelesen.
Man nimmt an, dass sich entweder bereits vor der Aufnahme des Proteobacteriums, spätestens aber im Zuge der Endosymbiose im Archaeon eine schützende Membran um seine eigene Erbsubstanz herum gebildet hat. Diese hat den Einbau von Fremd-DNA ins eigene Genom besser kontrollierbar gemacht. Vor allem aber hat sie einen Ort geschaffen, an dem die Erbsubstanz, die DNA, gespeichert wird. Hier wird ihre Information auch in RNA übersetzt, die anschließend verarbeitet und aus dem Kern heraus ins Cytoplasma transportiert wird, wo die Proteinsynthese stattfindet.

Die zwei primären Endosymbiosen. Die Endosymbiose eines Proteobakteriums durch ein Archaeon führt zur Bildung des ersten Mitochondriums. In diesem Zusammenhang bildet sich auch eine Membran um die Erbsubstanz herum (→ Zellkern). Aus den entstandenen Ur-Eukaryoten entwickeln sich die Pilze und Tiere. In einer zweiten Symbiose wird ein Cyanobakterium aufgenommen (→ Chloroplasten), es entsteht der Vorläufer der Pflanzenzellen. Beide Endosymbiosen sind mit einem massiven Gentransfer vom Organellen- ins Wirtsgenom verbunden (violette Pfeile). Die ursprünglichen Zellwände der Endosymbionten sowie die umhüllenden Membransysteme des Wirts werden aufgelöst und durch die Membranen aus Fettsäureester-Lipiden ersetzt, die für moderne Eukaryoten typisch sind.
Weitere Zellorganellen…
Im Zuge der Evolution haben sich in der Eukaryotenzelle viele weitere Organellen herausgebildet, die es in Prokaryoten und Archaeen nicht gibt. In diesen Membransystemen finden Stoffwechselprozesse statt und es werden Vesikel abgeschnürt, die Materialien an andere Orte innerhalb der Zelle oder nach draußen transportieren.
Prominente Beispiele sind der Golgi-Apparat und das unmittelbar an die Kernmembran anschließende und mit ihr verbundene endoplasmatische Retikulum. Daneben gibt es zahlreiche weitere Kompartimente, wie beispielsweise Lysosomen und Peroxysomen, in denen jeweils spezielle Stoffwechselreaktionen ablaufen.


Prokaryot und Eukaryot. Quellen: Prokaryot (oben): LadyofHats, german text by NEUROtiker – Derivative work of Image:Average prokaryote cell- en.svg by LadyofHats, Gemeinfrei, Link. Eukaryot (unten): Von Translation by Muellercrtp – Image:Plant cell structure svg.svg from Mariana Ruiz Villarreal. LadyofHats, Gemeinfrei, Link)
{kind=link}
Unter diesem Link https://www.thinglink.com/scene/565728773106827265 finden sich zwei sehr schöne dreidimensionale und interaktive Darstellungen von Pro- und Eukaryotenzellen.
Diese große innenliegende Membranfläche, insbesondere der Mitochondrien, ermöglicht den eukarotischen Zellen, sehr viel größer zu werden als es die prokaryotischen Zellen können.
Zellgröße
Die Größe, die eine Zelle erreichen kann, wird durch die Nährstoffversorgung sowohl von außen in die Zelle hinein als auch innerhalb der Zelle bestimmt. Entscheidend dafür ist das Verhältnis ihrer Oberfläche zum Volumen.
Bacteria und Archaea bleiben immer klein, sie sind durchschnittlich zwischen 1 und 10 µm groß. Die Nährstoffe werden innerhalb der Zelle durch Diffusion, also passives „Treiben“ im Zellplasma, verteilt.
Eukaryotische Zellen haben durch ihre Organellen zum einen ihre stoffwechselaktive Oberfläche drastisch vergößert – man denke nur an die unzähligen, der Energiegewinnung dienenden Membranauffaltungen in einem einzigen Mitochondrium! Zum anderen besitzen sie mit ihrem Cytoskelett, an dem entlang Vesikel mit allerlei Inhalt zwischen den Organellen entlang wandern können, ein effektives Transportsystem zum Verteilen von Nährstoffen, Signalmolekülen, Proteinen verschiedenster Art und Abfallprodukten innerhalb der Zelle. Eukaryotische Zellen können daher sehr viel größer werden als prokaryotische: Ihr Durchmesser reicht von 5 µm bei Hefezellen bis zu 1 mm bei Riesenamöben und Eizellen. Manche eukaryotische Zellen werden noch deutlich größer, etwa Nervenzellen des Menschen, deren Fortsätze bis zu 1 m lang werden können. Einzelne Zellen können auch zu sog. Synzytien verschmelzen und dadurch sehr groß oder langgestreckt werden (z.B. unsere Muskelzellen). Auch vielkernige Pflanzenzellen, z.B. Valonia untricularis, weisen Volumina von über 1 ml auf. Einige Armleuchteralgen können über 10 cm lange Zellschläuche ausbilden.
…und ein neuer Endosymbiont
Nachdem durch die oben beschriebene, erste Endosymbiose der Ur-Eukaryot (mit Mitochondrien, intrazellulärem Membransystem und Zellkern) entstanden ist, kam es im Lauf der Evolution zu einer zweiten Endosymbiose. Bei dieser nahm der Ur-Eukaryot ein Cyanobakterium in sich auf. Dieses bildete den Vorläufer der heutigen Chloroplasten und anderer Plastiden. Damit war der erste Vorläufer der heutigen Pflanzenzellen entstanden. Auch die Mehrzahl der Gene des Ur-Plastiden wurde mit der Zeit in den Zellkern verlagert und ins Kerngenom integriert. Der Gentransfer im Zuge der Endosymbiosen hat zur Ausbildung von Chromosomen geführt, die noch heute alle eukaryotischen Genome kennzeichnen.
Vergleiche von DNA- und rRNA-Sequenzen heutiger Eukaryoten zeigen, dass die beiden primären Endosymbiose-Ereignisse aller Wahrscheinlichkeit nach jeweils nur einmal im Laufe der Evolution stattgefunden haben. Alle heutigen Mitochondrien und alle heutigen Plastiden gehen also auf jeweils einen einzigen gemeinsamen Vorläufer zurück.
Im weiteren Verlauf der Evolution kam es zur Endosymbiose von Zellen, die bereits Plastiden enthielten (sekundäre Endosymbiosen). Beispiele dafür finden sich bei den Euglenophyta und Chlorarachniophyta. Auch tertiäre Endosymbiosen fanden statt, bei denen bereits sekundäre Plastiden besitzende Haptomonaden von Dinoflagellaten aufgenommen wurden. Auf diese Weise entstanden Plastiden-Varianten mit drei oder noch mehr Membranen.
Woher wissen wir, wann diese Ereignisse stattgefunden haben?
Den ältesten sicheren Hinweis auf Eukaryoten bilden heute Mikrofossilien, die auf 1,5 Milliarden Jahre datiert werden.
Biogeochemische Spuren bestimmter, für Eukaryoten typischer Fettmoleküle in deutlich älteren Schichten, die eine frühere Entstehung nahegelegt hätten, haben sich als Verunreinigungen erwiesen. [MM]