







375 Millionen Jahre vor heute
![]()
Zeitalter: Paläozoikum / Devon
 Mehrfache schnelle Veränderungen des Klimas und ein geringerer Sauerstoffgehalt im Wasser führen zum dritten großen Aussterben. Mit dazu bei trägt auch ein starker Anstieg des Vulkanismus, der vermutlich durch große Kontinentalverschiebungen hervorgerufen wird. Drei Viertel aller im Wasser lebenden Arten sind davon betroffen. Die Amphibien – Wirbeltiere, die Luft atmen und an Land leben können – erschließen sich das Land als neuen Lebensraum. Auch die ersten Quastenflosser treten auf – es gibt sie bis heute.
Mehrfache schnelle Veränderungen des Klimas und ein geringerer Sauerstoffgehalt im Wasser führen zum dritten großen Aussterben. Mit dazu bei trägt auch ein starker Anstieg des Vulkanismus, der vermutlich durch große Kontinentalverschiebungen hervorgerufen wird. Drei Viertel aller im Wasser lebenden Arten sind davon betroffen. Die Amphibien – Wirbeltiere, die Luft atmen und an Land leben können – erschließen sich das Land als neuen Lebensraum. Auch die ersten Quastenflosser treten auf – es gibt sie bis heute.
Dargestellt ist ein Tiktaalik roseae.

Flache warme Meere bedecken Teile der Kontinente. Das Klima ist zunächst warm, ähnlich wie heute in Europa. Später kühlen die Polargebiete allmählich ab. Besonders Südamerika, das nahe am Südpol liegt, ist davon betroffen.
Diese Infos in einfacher Sprache auf evokids.de.
Diese Station im Hörspiel:
Etwa 100 Mio. Jahre nach den Pflanzen, im Oberen Devon vor mehr als 390 Mio. Jahren, begannen auch die Wirbeltiere, das Land als Lebensraum zu erobern. Zuvor hatten bereits Würmer und Gliederfüßler das Festland erobert. Über die Gründe oder Zwänge für diese Entwicklung lässt sich nur spekulieren. Aus evolutionärer Sicht ist klar, dass die Umbildung der Extremitäten zumindest so weit fortgeschritten war, dass eine Fortbewegung auf dem Land möglich war. Wahrscheinlich hat die Entwicklung dort stattgefunden, wo die aquatischen Lebensräume der Tiere häufig trockenfielen und die Anpassung an eine zumindest teilweise terrestrische Lebensweise Vorteile bot. Aber auch eine Erweiterung oder Veränderung der bevorzugten Nahrung kann ein Selektionsvorteil für die Besiedlung des Festlandes gewesen sein.
Von Flossen zu Extremitäten
Als Ausgangspunkt dieser Entwicklung wurde von Anfang an die Ordnung der Quastenflosser (Crossopterygiformes) gesehen, deren heutige und fossile Vertreter vom anatomischen Standpunkt am ehesten für das Leben auf dem Trockenen geeignet erschienen. Mittlerweile haben jedoch molekulargenetische Untersuchungen gezeigt, dass Amphibien, Reptilien, Vögel und Säugetiere näher mit den heute noch vor bzw. in Afrika, Südamerika und Australien lebenden Lungenfischen (Dipnoi) verwandt sind und der Landgang der Wirbeltiere deshalb möglicherweise durch diese Unterklasse der Fleischflosser erfolgte.

Australischer Lungenfisch (Neoceratodus forsteri). (Urheber: Vassil – Lizenz:gemeinfrei (CC0 1.0) https://www.fischlexikon.eu/fischlexikon/fische-suchen.php?fisch_id=0000001522#)
Ziemlich gesichert ist, dass sich die Beine der zukünftigen Landwirbeltiere bereits im Wasser entwickelt haben. An einem gut erhaltenen Fossil eines Fleischflossers (Elpistostege) ist erkennbar, dass die Flossen bereits zu den Extremitäten der späteren Landwirbeltiere umgebildet wurden, obwohl es sich bei diesem Tier noch eindeutig um einen Fisch handelt. Trotzdem ähnelt das Skelett schon stark frühen Landwirbeltieren wie Ichthyostega oder Acanthostega, auch wegen der fehlenden unpaarigen Rücken- und Afterflosse, die sonst typisch für Fische ist. Sie unterscheiden sich jedoch im Knochenaufbau und in der Embryonalentwicklung, so dass ein evolutionärer Übergang aus den Einzelelementen schwer erklärbar ist. So bleiben bis heute Fragen bei den komplizierten phänotypischen Umbauten der Fischflossenskelettteile zu Elementen von Hand bzw. Fuß offen.

Ichthyostega, das vermutlich erste Landwirbeltier: Schädelrekonstruktion anhand von Fossilfunden (links) und Modell (rechts). Von den Extremitäten sind lediglich die hinteren fossil überliefert; sie sind paddelförmig gebaut. (Quelle: Schädel: FunkMonk, https://commons.wikimedia.org/wiki/File:Ichthyostega_skull.jpg; Modell: Dr. Günter Bechly, https://commons.wikimedia.org/wiki/File:Ichthyostega_model.jpg)
{kind=link}
Zu den Elpistostegalia gehört auch der auf dem Schild gezeigte Tiktaalik. Auch dessen Schädelknochen weisen darauf hin, dass es sich hier um eine Mosaikform zwischen Fisch und frühem Landwirbeltier handelt. Während Flossenstrahlen, Unterkiefer und Gaumen noch eindeutig denen von Fischen entsprechen, verfügen die Flossenstiele bereits über Ellenbogen und Handgelenk. Auch der Beckengürtel weist ebenso wie der Schädel bereits mehrere Merkmale auf, die später für die landlebenden Wirbeltiere typisch werden. Im Gegensatz zu den echten Fischen weist der Schädel von Tiktaalik je ein paariges Stirn- und Scheitelbein auf.

Fossil von Tiktaalik roseae im Königlich-Belgischen Institut für Naturwissenschaften. (Quelle: Esv – Eduard Solà und Petter Bøckman https://commons.wikimedia.org/wiki/File:Tiktaalik_belgium_II.jpg)
Voraussetzungen für das Leben an Land
Als erstes ausschließlich auf dem Land lebendes Tier wird Ichthyostega genannt, zumindest ist es das erste landlebende Tier, von dem wir Fossilfunde haben. Allerdings weist Ichthyostega ungewöhnlich viele Merkmale auf, die weder bei den Fischen, noch bei den nachfolgenden Landtieren zu finden sind, sodass angenommen werden darf, dass künftige Fossilfunde den Landgang der Wirbeltiere besser beschreiben können.
Die Fähigkeit, Luftsauerstoff zu atmen, findet sich bereits bei den frühen Lungenfischen, der Umbau der äußeren Hautschichten erfolgte erst nach dem Landgang.
Fortbewegung
Interessanterweise schwimmen Quastenflosser in einer Art „Kreuzgang“, ohne je an Land gelebt zu haben. Das bedeutet, rechtes Vorderbein und linkes Hinterbein, bzw. linkes Vorderbein und rechtes Hinterbein, werden mehr oder weniger gleichzeitig bewegt, bzw. vom Boden abgehoben und wieder aufgesetzt. Diese Fortbewegungsart ist auch bei Reptilien wie den Krokodilen zu beobachten und weist darauf hin, dass die Fortbewegung an Land von den Tieren nicht erst erlernt werden musste.
Anpassung der Fortpflanzung
Während die Amphibien zur Fortpflanzung noch auf Wasser angewiesen sind, in dem sie ihre Eier ablegen und die Tiere zumindest ihr Jugendstadium verbringen, mussten landlebende Wirbeltiere einen alternativen Mechanismus für die Fortpflanzung entwickeln. Sich komplett außerhalb des Wassers fortpflanzen zu können, hatte für frühe Landwirbeltiere das Potenzial, neue Lebensräume erschließen und so der Konkurrenz anderer Arten ausweichen zu können. Mit Entwicklung des Amnioten-Eies, wie es heute bei Reptilien, Vögeln und Kloakentieren vorkommt, konnte dieses Potenzial genutzt werden. Die Landwirbeltiere waren nunmehr in der Lage, sich auch in sehr trockene Regionen auszubreiten und die Kontinente flächendeckend zu besiedeln.
Kloakentiere (Monotremata) sind Säugetiere, die noch keinen lebenden Nachwuchs zur Welt bringen, sondern noch Eier legen. Heute noch lebende Beispiele dieser Ursäuger sind Ameisenigel und Schnabeltiere. Beide Gruppen kommen heute noch in Australien und Neuguinea vor und sind stark bedroht.

Ameisenigel (Kurzschnabeligel) und Schnabeltier als Beispiele für heute noch lebende Kloakentiere (Ursäuger). (Quellen: Ameisenigel: fir0002 | flagstaffotos.com.au, https://commons.wikimedia.org/wiki/File:Wild_shortbeak_echidna.jpg, Lizenz CC BY-NC; Schnabeltier: Stefan Kraft, https://commons.wikimedia.org/wiki/File:Platypus.jpg)
{kind=link}
Das Amnioten-Ei: Entwicklung im Wasser – auch auf dem Trockenen
Im Amnioten-Ei wächst der Embryo in einer von Flüssigkeit (Fruchtwasser) gefüllten Höhle (Amnionhöhle, Fruchtblase) heran. Diese ist völlig von einer Haut, dem Amnion, umschlossen und i.d.R. noch von weiteren Membranen bzw. Schalen umhüllt. Im Gegensatz zu den Amphibien, deren Entwicklung noch ein wasserlebendes Larvenstadium umfasst, reifen die Amnioten komplett in diesem Ei heran.
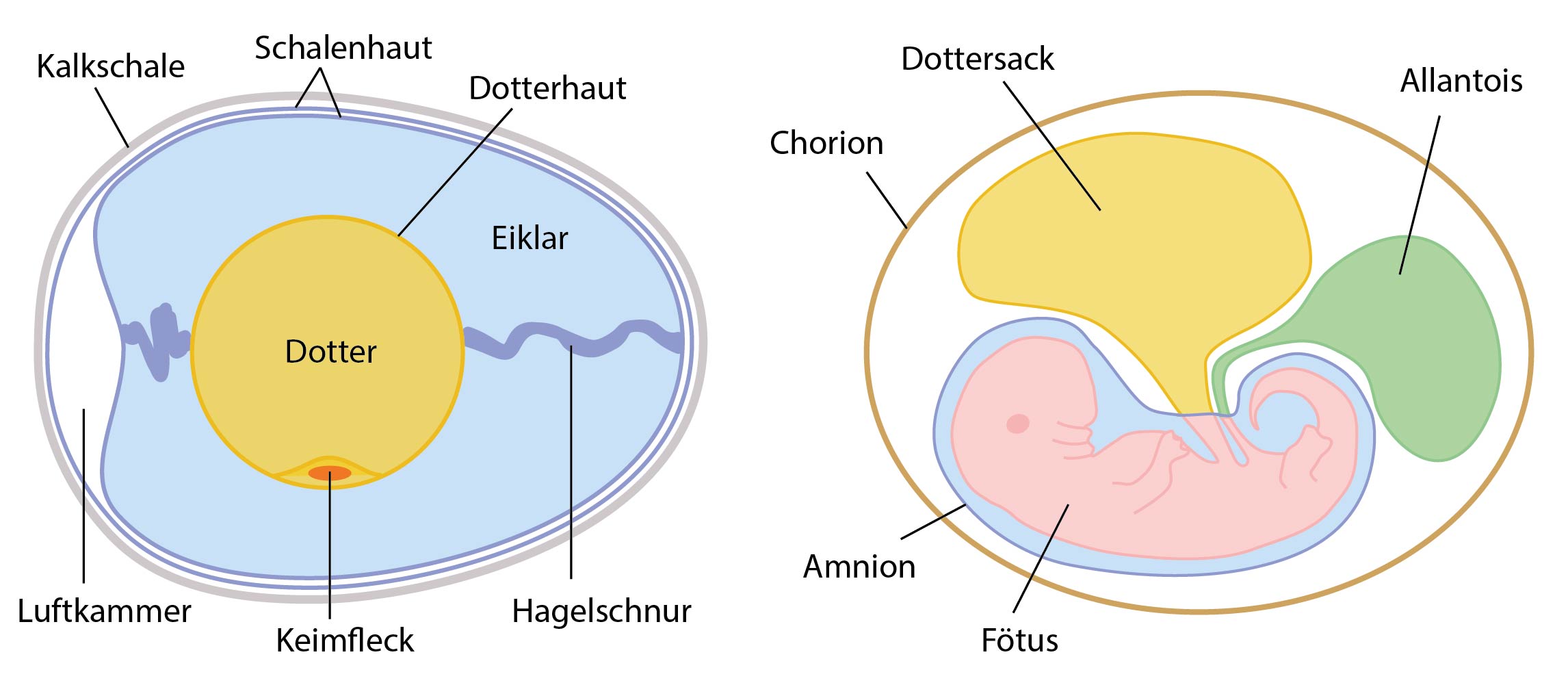
Amnioten-Ei: Bau am Beispiel eines unbebrüteten Hühnereies (links), und in einem Ei heranwachsender Embryo (rechts).
Mit seinem Wasserreservoir, dem Nährstoffvorrat und der schützenden Hülle fungiert das Ei als autonome Überlebenskapsel für den Embryo, dadurch wird ein Fötusstadium überhaupt erst möglich. Das Fruchtwasser schützt den Embryo unter anderem vor Austrocknung. Dottersack und Allantois sind mit intraembryonalen Räumen verbunden. Über den Dottersack werden dem Fötus Nährstoffe zugeführt, die Allantois nimmt stickstoffhaltige Stoffwechselprodukte des Embryos, also quasi den fetalen Harn, auf und wirkt beim Gasaustausch mit.
Schutz gegen Austrocknung
Ähnlich wie schon früher die Pflanzen bei ihrem Landgang haben auch die Wirbeltiere einen Schutz ihrer Außenhaut gegen Austrocknung entwickelt. Sie besteht meistens aus Faserproteinen (Keratin), auch Horn genannt. Diese Eiweiße sind wasserunlöslich und schwer abbaubar, weshalb sie sowohl als Verdunstungssperre, aber auch als mechanischer Schutz gegen Verletzungen dienen.
Entwicklung der Wirbeltiere auf dem Land
An der Basis der Entwicklung zu Landwirbeltieren standen die Seymouria. Hierbei handelt es sich um eine Mosaikform, die Merkmale von Amphibien und Reptilien vereinigt und die sich im frühen Perm entwickelten. Zu den Reptilienmerkmalen rechnet man vor allem das Skelett. Die Ursprünglichkeit des Schädels wird durch die Tatsache unterstrichen, dass noch keine Schläfenfenster vorhanden waren. Die fehlenden Schläfenfenster sind ein Charakteristika für die Gruppe der Anapsiden, die auch als Schläfengrubenlose bezeichnet werden.
Aus einer Schwestergruppe der Seymuoriamorpha, den Diadectomorpha, entwickelte sich bereits früh die Linie der Chelonia, die heute nur noch durch die Suppenschildkröte (Chelonia mydas), vertreten ist. Eine weitere frühe Abspaltung ergab sich durch die Entwicklung der oben beschriebenen Ichthyosaurier und der Plesiosaurier. Beide Linien haben sich aus den Stammreptilien entwickelt.
Im Zeitalter der Trias begann die Ausbreitung der ersten Reptilien mit zwei Schläfenfenstern (Diapside), die sich aus den Eosuchiern entwickelten. Von den Eosuchiern lassen sich die Archosaurier und Lepidosaurier sowie die Avicephala ableiten. Aus den Eosuchiern entwickelten sich bereits im Perm auch schildkrötenähnliche Reptilien, die ihre äußeren Merkmale bis heute bewahrt haben.
Aus den urtümlichen Lepidosauria entstanden die Schuppenkriechtiere, insbesondere die Echsen und die Schlangen. Sowohl Echsen als auch Schlangen zeichnen sich durch einen diapsiden Schädeltypus aus, der durch die Reduktion des Jochbogens eine weitgehende Abwandlung erfahren hat. Eine weitere Gruppe, die Thecodontier, entwickelte sich vom Ende des Perm bis ins Trias. Die Thecodontier und davon abgeleitete Formen werden heute als Archosaurier zusammengefasst. Im späteren Verlauf der Entwicklung, vom Jura bis zur Kreide, entwickelten sich aus den Thecodontiern auch die allen bekannten Dinosaurier sowie die Pterosaurier. Eine weitere Linie führt zu den Vögeln. Die Entwicklung der Vögel begann aber wahrscheinlich erst im Jura. [KHB]