







2500 Millionen Jahre
oder 610 Meter vor heute
Zeitalter: Proterozoikum / Siderium
 Zellen entwickeln die Fähigkeit, aus Sonnenlicht immer effizienter Energie zu gewinnen. Der entstehende Sauerstoff führt über Millionen von Jahren lediglich zur Bildung von wasserunlöslichen Metallsalzen in den Ozeanen. Es entstehen die mächtigen Bändererzschichten, die wir noch heute abbauen.
Zellen entwickeln die Fähigkeit, aus Sonnenlicht immer effizienter Energie zu gewinnen. Der entstehende Sauerstoff führt über Millionen von Jahren lediglich zur Bildung von wasserunlöslichen Metallsalzen in den Ozeanen. Es entstehen die mächtigen Bändererzschichten, die wir noch heute abbauen.
Die Eisenkonzentration im Wasser sinkt ab, so dass freier Sauerstoff auftritt. Der ist allerdings giftig: Das erste große Aussterben („Große Sauerstoffkatastrophe“) erfasst die meisten Arten. Organismen, die in sauerstofffreier Umgebung leben, bleiben unbehelligt. Neue Arten, die Sauerstoff vertragen oder damit sogar viel Energie gewinnen, breiten sich aus.
 Erster Sauerstoff gelangt in die Atmosphäre, sein Anteil steigt stetig. Parallel dazu sinkt der Kohlendioxid-Gehalt. Die Erde ist überwiegend von Ozeanen bedeckt. Nur vereinzelt überragen Vulkankegel oder angehobene Erdschollen (Kratone) die Wasseroberfläche.
Erster Sauerstoff gelangt in die Atmosphäre, sein Anteil steigt stetig. Parallel dazu sinkt der Kohlendioxid-Gehalt. Die Erde ist überwiegend von Ozeanen bedeckt. Nur vereinzelt überragen Vulkankegel oder angehobene Erdschollen (Kratone) die Wasseroberfläche.
Diese Infos in einfacher Sprache auf evokids.de.
Die Photosynthese ist die wichtigste Reaktion für das Leben auf der Erde in seiner heutigen Form. Ihre Bedeutung kann gar nicht hoch genug geschätzt werden. Denn sie ermöglicht es allen tierischen Lebewesen und vielen Bakterien und Pilzen, und natürlich auch den photosynthese-treibenden Organismen selbst, die Energie der Sonne als Lebensgrundlage zu nutzen. Die allermeisten heute lebenden Organismen sind für ihren Stoffwechsel auf die Produkte der Photosynthese treibenden Organismen angewiesen.
Heute findet die Photosynthese in extrem großem Umfang statt, sie wird überwiegend von den grünen Landpflanzen geleistet. Diese binden alljährlich etwa 120 Milliarden Tonnen CO2, von denen etwa die Hälfte in Form von Biomasse fixiert wird (die andere Hälfte des fixierten CO2 wird im Zuge des Stoffwechsels der Pflanzen wieder frei und in die Atmosphäre abgegeben). Hinzu kommen pro Jahr etwa 55 Milliarden Tonnen Biomasse, die durch marine Organismen photosynthetisiert wird.
Vor 2,5 Milliarden Jahren gab es natürlich noch keine höheren Pflanzen, noch nicht einmal Einzeller mit Zellkern waren entwickelt. In dieser Frühzeit des Lebens wurde die oxygene (Sauerstoff-freisetzende) Photosynthese von Cyanobakterien „erfunden“, die zusammen mit mineralhaltigen Sedimenten Matten auf Gesteinen im Ozean bildeten. Reste davon sind in Form der Stromatolithen noch heute zu sehen.
Was passiert bei der Photosynthese, und warum ist sie so wichtig?
Cyanobakterien haben die Fähigkeit entwickelt, die Energie des Sonnenlichtes zu nutzen, um das in der Umgebung vorhandene CO2 in Form energiereicher Verbindungen (Zucker!) zu fixieren. Diese energiereichen Verbindungen dienen wiederum den allermeisten anderen Organismen, insbesondere den Tieren und auch uns Menschen, als Energiequelle: Wir nehmen sie als Nahrung auf und „verbrennen“ sie (mit Hilfe des bei der Photosynthese als Nebenprodukt entstehenden Sauerstoffs) wieder zu CO2. Die dabei freiwerdende Energie treibt all unsere Stoffwechselprozesse an.
Dafür gibt es übrigens Fachbegriffe: Cyanobakterien und alle höheren Pflanzen leben „photoautotroph“ (das bedeutet: sie gewinnen ihre Energie aus Licht und bauen ihre Biomasse selbst aus CO2 auf), alle Tiere leben „chemoheterotroph“ (sie gewinnen ihre Energie aus chemischen Reaktionen und bauen ihre Biomasse, indem sie andere Biomasse umwandeln).

Stoff- und Energiekreislauf auf der Erde. Aus energiearmen anorganischen Stoffen werden im Zuge der Photosynthese energiereiche organische Stoffe erzeugt – unter Nutzung der Sonnenenergie. Diese energiereichen Stoffe dienen wiederum anderen Organismen als Nahrung und Energiespender.
Für alle, die es genauer wissen wollen:
Die Photosynthese läuft in zwei Phasen ab. In der sogenannten Primärreaktion (auch „Lichtreaktion“ genannt) wird mithilfe von großen Proteinkomplexen in Membranen, den sogenannten Photosystemen, die Energie aus dem Sonnenlicht in universellen biologischen Energieträger-Molekülen gespeichert: ATP ist vielen ein Begriff, aber auch sogenannte Reduktionsäquivalente [H] sind Energieträger.
- Einfache Organismen nutzen nur ein Photosystem, das jedoch nicht genug Energie liefert, um Elektronen aus Wasser herauszulösen. Statt Wasser verwendet es beispielsweise H2S oder einfache organische Substanzen als Elektronenquelle. Bei diesen Formen der Photosynthese, die bereits von Vorläufern der Cyanobakterien betrieben wurde und auch heute noch vielen Bakterienarten zur Energiegewinnung dient, wird kein Sauerstoff freigesetzt. Die Energie dieses einen Photosystems genügt lediglich zur Bildung von Reduktionsäquivalenten, die für weitere Biosynthesen verwendet werden können.
- Bei der voll evolvierten, oxygenen Photosynthese der Cyanobakterien und heutigen Landpflanzen sind zwei Photosysteme hintereinandergeschaltet – quasi wie bei einem Motor mit Turbolader oder einem Zug mit zwei Loks. Erst das zweite Photosystem liefert genug Energie, um Wasser in Wasserstoff und Sauerstoff zu spalten. Der Sauerstoff diffundiert weg, Wasserstoff-Ionen (H+) werden jedoch angereichert und zur Bildung von ATP genutzt.
In der Sekundärreaktion (sie ist nicht lichtabhängig und wird daher häufig etwas irreführend als „Dunkelreaktion“ bezeichnet), werden die zuvor erzeugten Energieträger dazu verwendet, mithilfe eines speziellen Enzyms ein CO2-Molekül an einen C5-Zucker zu ketten, wobei dieser zugleich in zwei C3-Zucker gespalten wird. Aus diesen können dann wiederum weitere Zuckermoleküle mit 3, 4, 5, 6 oder 7 Kohlenstoffmolekülen hergestellt werden, und aus diesen alle von dem Organismus benötigten Bausteine – weitere Zucker, Fette, Eiweiße.
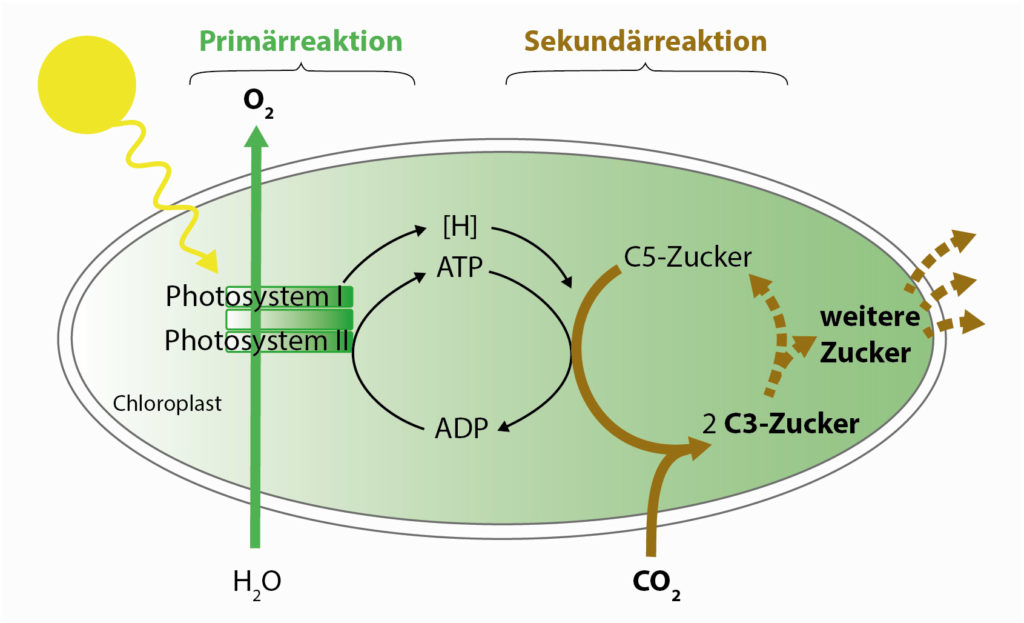
Die zwei Schritte der Photosynthese: In der Primärreaktion wird die Energie aus dem Sonnenlicht in biochemischen Energieträgern (ATP und [H]) gespeichert. In der Sekundärreaktion wird diese Energie zur Bindung eines CO2-Moleküls an ein 5er-Zuckermolekül genutzt. Aus den entstehenden 3er-Zuckern wiederum können viele weitere Lebensmoleküle gebildet werden.
Leben ohne und mit Photosynthese
Nun gab es bekanntlich auch vor der Entwicklung der Photosynthese bereits Leben auf der Erde. Diese ganz frühen Lebensformen nutzten die relativ geringen Energiegradienten einfacher physikochemischer Reaktionen zum Antrieb ihres Stoffwechsels (in Fachsprache: sie lebten chemoautotroph). Die Fähigkeit zur Photosynthese, also zur Nutzung der im Sonnenlicht enthaltenen Energie, brachte ihren „Erfindern“ ungeheure Vorteile gegenüber diesen frühen Lebensformen.
Und Leben, wie es heute auf der Erde existiert, wäre ohne die oxygene Photosynthese überhaupt nicht möglich.
- Die photosynthetisch gebildeten Moleküle dienen allen höheren Organismen als Nahrung. Sie nutzen sie als Bauelemente, und mit der darin enthaltenen Energie treiben sie ihren Stoffwechsel an.
- Ohne Photosynthese würden allerdings nicht nur die organischen Bausteine fehlen, sondern auch der Sauerstoff für deren effizienten Abbau.
Wir Menschen benötigen für unseren Stoffwechsel bekanntlich Sauerstoff – wir sind aerobe Organismen. Ebenso alle Tiere und Pflanzen. Vielen Bakterien und Mikroorganismen (sogenannten Anaerobiern) gelingt ihr Stoffwechsel jedoch auch ohne Sauerstoff, sie ersetzen ihn beispielsweise durch Stickstoff oder Schwefel. - Und nicht zuletzt wäre die Entwicklung des heute so vielgestaltigen Lebens ohne die schützende Ozonschicht in der äußeren Atmosphäre gar nicht möglich gewesen: Das Ozon (O3) absorbiert ein Großteil der für alle Lebewesen schädlichen UV-Strahlung, und es existiert selbstverständlich auch erst, seit es in der Atmosphäre freien Sauerstoff gibt.
- Im Übrigen sind auch alle fossilen Energieträger, die wir heute zum „Antrieb“ unserer nicht biologisch bewegten Prozesse oder zur Wärmegewinnung nutzen (z.B. Braunkohle, Steinkohle, Erdöl), Folgeprodukte der Photosynthese.
Allerdings blieb die Photosynthese, wie überhaupt das gesamte Leben, zunächst über mehr als 2 Milliarden Jahre auf die Ozeane beschränkt, betrieben von frei im Wasser schwimmenden oder auf Gesteinen im Flachwasser fixierten Cyanobakterien. Erst mit der Ausbreitung früher pflanzlicher Lebensformen über die Wasserlinie hinaus (zunächst noch in Form ausgedehnter Matten auf Steinen im Gezeitensaum und sehr viel später durch den Landgang der ersten Pflanzen) gelangte das Leben ans Land, und der photosynthetische Gasaustausch fand direkt in der Atmosphäre statt.
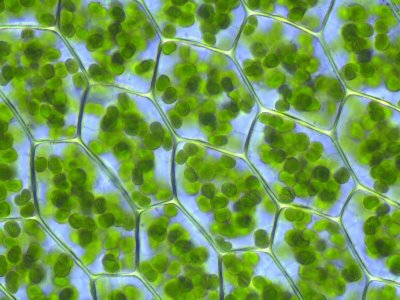
Chloroplasten im Blatt eines Laubmooses: Chloroplasten sind die photosynthetisch aktiven Zellorganellen heutiger Algen und Pflanzen. Sie sind durch Endosymbiose von Cyanobakterien entstanden. (Foto: Kristian Peters, aus wikipedia, Artikel “Photosynthese”. Lizenz: GNU FDL 1.2.)
Woher wissen wir, wann sich die Photosynthese entwickelt hat?
Vorweg: Genau wissen wir das nicht, sondern das wird nach wie vor intensiv beforscht. Vielleicht ist die Photosynthese schon vor 3,8 Milliarden Jahren entstanden, wahrscheinlich vor über 3 Milliarden Jahren, und mit ziemlicher Sicherheit gab es sie spätestens vor 2,7 Milliarden Jahren.
Fossilfunde von Cyanobakterien
Für eine sehr frühe Entstehung sprechen fossile Reste der bereits erwähnten Stromatolithen, die wir bereits in 3,4 Milliarden Jahre alten Schichten finden. Ganz sicher gegeben hat es sie vor 2,7 Mrd Jahren – so alt sind die ältesten fossilen Cyanobakterien, die man bisher gefunden hat.
Nachweisbare Folgen der steigenden Sauerstoffkonzentration
Fast wichtiger als diese Fossilien sind für die zeitliche Einordnung die Veränderungen, die der im Zuge der oxygenen Photosynthese gebildete Sauerstoff in seiner Umgebung hervorgerufen hat und die noch heute nachweisbar sind.
Bis vor 3 Milliarden Jahren gab es Sauerstoff in der Erdatmosphäre nur in Spuren. Er entstand, wenn durch UV-Strahlung in den äußeren Schichten der Atmosphäre Wasser gespalten wurde. Der dabei ebenfalls entstehende Wasserstoff ist ins Weltall verschwunden, der (schwerere) Sauerstoff wurde durch die Erdanziehung in der Atmosphäre gehalten.
Mit der Entwicklung der ersten Photosynthese betreibenden Organismen nahm der Sauerstoffgehalt der Atmosphäre langsam zu. Nachweisbar ist das anhand von verschiedenen Chrom-Isotopen, die unterschiedlich leicht oxidiert werden. 53Cr wird sehr viel leichter oxidiert als 52Cr. Während oxidiertes Chrom leicht aus dem Gestein ausgewaschen wird, bleibt nicht oxidiertes Chrom im Gestein gebunden. Die Analyse von 3 bis 2,4 Milliarden Jahren alten Gesteinsschichten zeigt, dass damals der Anteil von 53Cr im Festlandsgestein gesunken ist, während er in ozeanischem Sedimentgestein angestiegen ist – ein Hinweis auf die steigende O2-Konzentration in der Atmosphäre, die zur Oxidation des 53Cr und zu seiner Auswaschung geführt hat.
Der Sauerstoffgehalt der Ozeane veränderte sich, verglichen mit dem der Atmosphäre, nur langsam: Der Sauerstoff hat zunächst das damals in großer Menge in den Ozeanen gelöste Fe2+ oxidiert. Dabei entstand unlösliches Fe3+ (in Form von Fe2O3 oder Eisenoxid), das sich als Rost am Meeresboden absetzte. Diese Eisen-Ablagerungen in etwa 2,7 Milliarden Jahre alten Gesteinsschichten sind noch heute als sogenannte Bändererze erhalten – inzwischen meist nicht mehr unter dem Meeresboden, sondern eingebettet in jene Gesteinsschichten, aus denen sie von uns Menschen abgebaut werden und nicht zuletzt die Basis unserer Industrialisierung bilden (über 90 % des wirtschaftliche genutzten Eisens stammen aus den vor 2,7 Milliarden Jahren abgelagerten Erzschichten!).

Bändererz mit typischer Schichtung aus Eisenoxid-Mineralen und kieseligen Sedimenten. Foto: André Karwath.
CC BY-SA 2.5Hinweise zur Weiternutzung; File:Black-band ironstone (aka).jpg; Erstellt: 26. August 2005
Standort: 51° 2′ 34,84″ N, 13° 45′ 26,67″ E
.jpg¶ms=51.043011_N_13.757408_E_&language=de){kind=link}
Sauerstoff – Gift und Lebensspender zugleich
Lange Zeit änderte sich die Konzentration an freiem Sauerstoff im Wasser und in der Atmosphäre nur langsam, weil er fast im selben Maße, wie er entstand, in Form von Metalloxiden wieder fixiert wurde. Als jedoch die Metalle, allen voran das Fe2+, verbraucht waren, stieg die Konzentration plötzlich extrem schnell.
Es ist sehr wahrscheinlich, dass die frühen, obligat anaeroben Organismen mit dem plötzlich in ihrer Umgebung vorhandenen Sauerstoff nicht zurechtkamen: Für nicht an O2 angepasste Lebensformen ist er hochgiftig. Er führt in deren Stoffwechsel zur Bildung hochreaktiver Peroxide, die wichtige strukturelle Bestandteile (Membranen, Eiweiße) beschädigen. Die meisten bis dahin vorhandenen anaeroben Lebensformen starben aus. Erst die Entwicklung von Peroxidasen – Enzymen, die schädliche Peroxide spalten können – ermöglichte es den Organismen, in Anwesenheit von Sauerstoff zu überleben.
Ein weiterer Faktor, der zu der „Großen Sauerstoffkatastrophe“ beigetragen hat, war die Oxidation von atmosphärischem Methan (CH4) zu CO2 und Wasser. Methan ist ein noch sehr viel stärkeres Treibhausgas als CO2. Die sinkende Methankonzentration war Mitauslöser einer extremen Eiszeit („Schneeball-Erde“), die 300 bis 400 Millionen Jahre lang andauerte und vermutlich ebenfalls viele der damals vorhandenen primitiven Lebensformen aussterben ließ.
Vor 2,2 Milliarden Jahren stieg der Sauerstoffgehalt der Atmosphäre noch einmal deutlich an (auf fast ein Fünftel des heutigen Gehalts, also auf etwa 3,8%). Man vermutet, dass das mit dem Ende der Vereisungsphase zusammenhängt: Durch Gletschererosion wurde Nachschub an Mineralien in die Ozeane gespült, und das durch die Vulkanausbrüche in die Atmosphäre getragene CO2 lieferte das nötige Material für eine massive Zunahme an photosynthetisch aktiven Organismen – eine erste weltweite „Algenblüte“ war die Folge. Zu dieser Zeit bedeckten ausgedehnte Biofilme die Steine im Flachwasserbereich. Mit einem Sauerstoffgehalt von 3 % wurde die Schwelle erreicht, die zur Ausbildung der Ozonschicht in der Stratosphäre führte. Diese schirmte die lebensfeindliche UV-Strahlung ab und ermöglichte eine Weiterentwicklung des Lebens und auch seine Ausbreitung an Land. Vor etwa 350 Millionen Jahren erreichte der Sauerstoffgehalt der Atmosphäre das heutige Niveau von 21%.
Die Entwicklung des aeroben Stoffwechsels, also von Enzymsystemen, die Biomoleküle mit Hilfe von Sauerstoff abbauen und damit sehr viel mehr Energie freisetzen (Zellatmung), als sich mit anaeroben Prozessen gewinnen lässt, ermöglichte schließlich die Evolution einer großen Vielfalt an höheren Lebensformen: Alle mehrzelligen Lebensformen der Erde sind auf Sauerstoff als Oxidationsmittel angewiesen (einzige Ausnahme ist ein erst vor wenigen Jahren im Bodensediment des Mittelmeeres entdeckter, weniger als einen Millimeter großer Organismus aus der Gruppe der Loricifera). Praktisch alle dauerhaft ohne Sauerstoff lebensfähigen Organismen sind also Einzeller – die meisten davon Bakterien und Archaeen, nur einige wenige sind eukaryote Einzeller, besitzen also auch einen Zellkern. [MM]